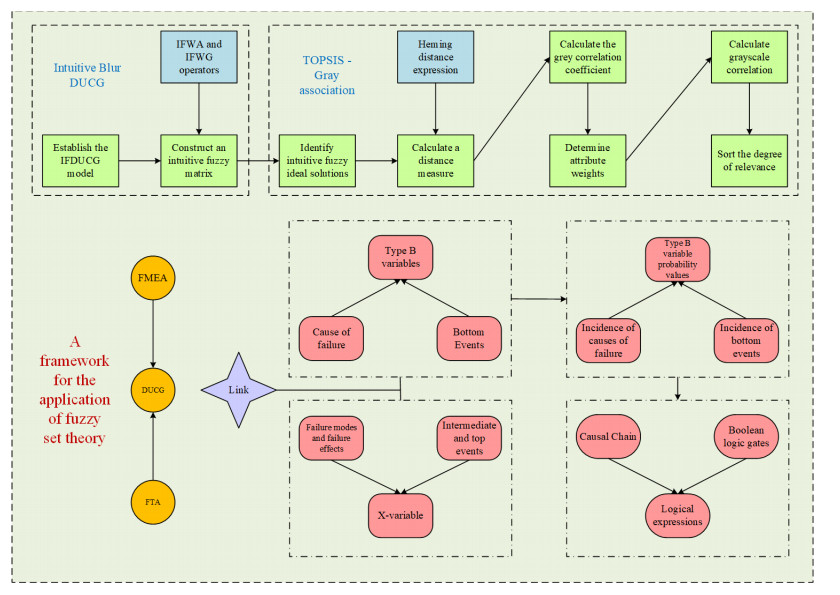
Induction motors have been widely used in industry, agriculture, transportation, national defense engineering, etc. Defects of the motors will not only cause the abnormal operation of production equipment but also cause the motor to run in a state of low energy efficiency before evolving into a fault shutdown. The former may lead to the suspension of the production process, while the latter may lead to additional energy loss. This paper studies a fuzzy rule-based expert system for this purpose and focuses on the analysis of many knowledge representation methods and reasoning techniques. The rotator fault of induction motors is analyzed and diagnosed by using this knowledge, and the diagnosis result is displayed. The simulation model can effectively simulate the broken rotator fault by changing the resistance value of the equivalent rotor winding. And the influence of the broken rotor bar fault on the motors is described, which provides a basis for the fault characteristics analysis. The simulation results show that the proposed method can realize fast fault diagnosis for rotators of induction motors.
Citation: Tangsheng Zhang, Hongying Zhi. A fuzzy set theory-based fast fault diagnosis approach for rotators of induction motors[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 9268-9287. doi: 10.3934/mbe.2023406
Induction motors have been widely used in industry, agriculture, transportation, national defense engineering, etc. Defects of the motors will not only cause the abnormal operation of production equipment but also cause the motor to run in a state of low energy efficiency before evolving into a fault shutdown. The former may lead to the suspension of the production process, while the latter may lead to additional energy loss. This paper studies a fuzzy rule-based expert system for this purpose and focuses on the analysis of many knowledge representation methods and reasoning techniques. The rotator fault of induction motors is analyzed and diagnosed by using this knowledge, and the diagnosis result is displayed. The simulation model can effectively simulate the broken rotator fault by changing the resistance value of the equivalent rotor winding. And the influence of the broken rotor bar fault on the motors is described, which provides a basis for the fault characteristics analysis. The simulation results show that the proposed method can realize fast fault diagnosis for rotators of induction motors.

| [1] |
Z. Guo, K. Yu, N. Kumar, W. Wei, S. Mumtaz, M. Guizani, Deep distributed learning-based POI recommendation under mobile edge networks, IEEE Internet Things J., 10 (2022), 303–317. https://doi.org/10.1109/JIOT.2022.3202628 doi: 10.1109/JIOT.2022.3202628

|
| [2] |
Y. Li, H. Ma, L. Wang, S. Mao, G. Wang, Optimized content caching and user association for edge computing in densely deployed heterogeneous networks, IEEE Trans. Mob. Comput., 21 (2020), 2130–2142. https://doi.org/10.1109/TMC.2020.3033563 doi: 10.1109/TMC.2020.3033563
|
| [3] |
Z. Guo, K. Yu, A. K. Bashir, D. Zhang, Y. D. Al-Otaibi, M. Guizani, Deep information fusion-driven POI scheduling for mobile social networks, IEEE Network, 36 (2022), 210–216. https://doi.org/10.1109/MNET.102.2100394 doi: 10.1109/MNET.102.2100394
|
| [4] |
L. Yang, Y. Li, S. X. Yang, Y. Lu, T. Guo, K. Yu, Generative adversarial learning for intelligent trust management in 6G wireless networks, IEEE Network, 36 (2022), 134–140. https://doi.org/10.1109/MNET.003.2100672 doi: 10.1109/MNET.003.2100672
|
| [5] |
Q. Zhang, K. Yu, Z. Guo, S. Garg, J. J. P. C. Rodrigues, M. M. Hassan, et al., Graph neural networks-driven traffic forecasting for connected internet of vehicles, IEEE Trans. Network Sci. Eng., 9 (2022), 3015–3027. https://doi.org/10.1109/TNSE.2021.3126830 doi: 10.1109/TNSE.2021.3126830
|
| [6] |
L. Zhao, Z. Bi, A. Hawbani, K. Yu, Y. Zhang, M. Guizani, ELITE: An intelligent digital twin-based hierarchical routing scheme for softwarized vehicular networks, IEEE Trans. Mob. Comput., 2022 (2022). https://doi.org/10.1109/TMC.2022.3179254 doi: 10.1109/TMC.2022.3179254
|
| [7] |
Y. Zhu, W. Zheng, Observer-based control for cyber-physical systems with DoS attacks via a cyclic switching strategy, IEEE Trans. Autom. Control, 65 (2019), 3714–3721. https://doi.org/10.1109/TAC.2019.2953210 doi: 10.1109/TAC.2019.2953210
|
| [8] |
L. Zhao, Z. Yin, K. Yu, X. Tang, L. Xu, Z. Guo, et al., A fuzzy logic based intelligent multi-attribute routing scheme for two-layered SDVNs, IEEE Trans. Netw. Serv. Manage., 2022 (2022). https://doi.org/10.1109/TNSM.2022.3202741 doi: 10.1109/TNSM.2022.3202741
|
| [9] |
L. Chen, Y. Zhu, C. K. Ahn, Adaptive neural network-based observer design for switched systems with quantized measurements, IEEE Trans. Neural Networks Learn. Syst., 2021 (2021), 1–14. https://doi.org/10.1109/TNNLS.2021.3131412 doi: 10.1109/TNNLS.2021.3131412
|
| [10] |
J. Zhang, Q. Yan, X. Zhu, K. Yu, Smart industrial IoT empowered crowd sensing for safety monitoring in coal mine, Digital Commun. Networks, 2022 (2022). https://doi.org/10.1016/j.dcan.2022.08.002 doi: 10.1016/j.dcan.2022.08.002
|
| [11] |
Z. Cai, X. Zheng, A private and efficient mechanism for data uploading in smart cyber-physical systems, IEEE Trans. Network Sci. Eng., 7 (2020), 766–775. https://doi.org/10.1109/TNSE.2018.2830307 doi: 10.1109/TNSE.2018.2830307
|
| [12] |
Z. Guo, K. Yu, A. Jolfaei, F. Ding, N. Zhang, Fuz-Spam: Label smoothing-based fuzzy detection of spammers in internet of things, IEEE Trans. Fuzzy Syst., 30 (2022), 4543–4554. https://doi.org/10.1109/TFUZZ.2021.3130311 doi: 10.1109/TFUZZ.2021.3130311
|
| [13] |
Z. Cai, X. Zheng, J. Yu, A differential-private framework for urban traffic flows estimation via taxi companies, IEEE Trans. Ind. Inf., 15 (2019), 6492–6499. https://doi.org/10.1109/TⅡ.2019.2911697 doi: 10.1109/TⅡ.2019.2911697
|
| [14] |
Z. Zhou, Y. Li, J. Li, K. Yu, G. Kou, M. Wang, et al., Gan-siamese network for cross-domain vehicle re-identification in intelligent transport systems, IEEE Trans. Network Sci. Eng., 2022 (2022), 1–12. https://doi.org/10.1109/TNSE.2022.3199919 doi: 10.1109/TNSE.2022.3199919
|
| [15] |
Z. Zhou, Y. Su, J. Li, K. Yu, Q. J. Wu, Z. Fu, et al., Secret-to-image reversible transformation for generative steganography, IEEE Trans. Dependable Secure Comput., 2022 (2022), 1–17. https://doi.org/10.1109/TDSC.2022.3217661 doi: 10.1109/TDSC.2022.3217661
|
| [16] |
S. Xia, Z. Yao, Y. Li, S. Mao, Online distributed offloading and computing resource management with energy harvesting for heterogeneous MEC-enabled IoT, IEEE Trans. Wireless Commun., 20 (2021), 6743–6757. https://doi.org/10.1109/TWC.2021.3076201 doi: 10.1109/TWC.2021.3076201
|
| [17] |
Z. Guo, Y. Shen, S. Wan, W. Shang, K. Yu, Hybrid intelligence-driven medical image recognition for remote patient diagnosis in internet of medical things, IEEE J. Biomed. Health Inf., 26 (2022), 5817–5828. https://doi.org/10.1109/JBHI.2021.3139541 doi: 10.1109/JBHI.2021.3139541
|
| [18] |
Z. Cai, Q. Chen, Latency-and-coverage aware data aggregation scheduling for multihop battery-free wireless networks, IEEE Trans. Wireless Commun., 20 (2021), 1770–1784. https://doi.org/10.1109/TWC.2020.3036408 doi: 10.1109/TWC.2020.3036408
|
| [19] |
C. Chen, Z. Liao, Y. Ju, C. He, K. Yu, S. Wan, Hierarchical domain-based multi-controller deployment strategy in SDN-enabled space-air-ground integrated network, IEEE Trans. Aerosp. Electron. Syst., 58 (2022), 4864–4879. https://doi.org/10.1109/TAES.2022.3199191 doi: 10.1109/TAES.2022.3199191
|
| [20] |
H. Jafari, J. Poshtan, Fault detection and isolation based on fuzzy‐integral fusion approach, IET Sci. Meas. Technol., 13 (2019), 296–302. https://doi.org/10.1049/iet-smt.2018.5005 doi: 10.1049/iet-smt.2018.5005
|
| [21] |
Y. Lu, L. Yang, S. X. Yang, Q. Hua, A. K. Sangaiah, T. Guo, et al., An intelligent deterministic scheduling method for ultralow latency communication in edge enabled industrial internet of things, IEEE Trans. Ind. Inf., 19 (2022), 1756–1767. https://doi.org/10.1109/TⅡ.2022.3186891 doi: 10.1109/TⅡ.2022.3186891
|
| [22] |
S. K. Gundewar, P. V. Kane, Condition monitoring and fault diagnosis of induction motor, J. Vib. Eng. Technol., 9 (2021), 643–674. https://doi.org/10.1007/s42417-020-00253-y doi: 10.1007/s42417-020-00253-y
|
| [23] |
A. Choudhary, D. Goyal, S. L. Shimi, A. Akula, Condition monitoring and fault diagnosis of induction motors: A review, Arch. Comput. Methods Eng., 26 (2019), 1221–1238. https://doi.org/10.1007/s11831-018-9286-z doi: 10.1007/s11831-018-9286-z
|
| [24] |
D. Fan, G. P. Jiang, Y. R. Song, Y. W. Li, G. Chen, Novel epidemic models on PSO-based networks, J. Theor. Biol., 477 (2019), 36–43. https://doi.org/10.1016/j.jtbi.2019.06.006 doi: 10.1016/j.jtbi.2019.06.006
|
| [25] |
I. Dilmi, A. Bouguerra, A. Djrioui, L. Chrifi-Alaoui, Interval type-2 fuzzy logic-second order sliding mode based fault detection and active fault-tolerant control of brushless DC motor, J. Eur. Syst. Automatisés, 54 (2021), 475–485. https://doi.org/10.18280/jesa.540311 doi: 10.18280/jesa.540311
|
| [26] |
O. E. Hassan, M. Amer, A. K. Abdelsalam, B. W. Williams, Induction motor broken rotor bar fault detection techniques based on fault signature analysis–a review, IET Electr. Power Appl., 12 (2018), 895–907. https://doi.org/10.1049/iet-epa.2018.0054 doi: 10.1049/iet-epa.2018.0054
|
| [27] | H. MERABET, T. Bahi, K. BEDOUD, D. DRICI, A fuzzy logic based approach for the monitoringof open switch fault in a SVM voltage sourceinverter fed induction motor drive, J. Autom. Syst. Eng., 12 (2018), 48–66. |
| [28] |
P. Kumar, A. S. Hati, Deep convolutional neural network based on adaptive gradient optimizer for fault detection in SCIM, ISA Trans., 111 (2021), 350–359. https://doi.org/10.1016/j.isatra.2020.10.052 doi: 10.1016/j.isatra.2020.10.052
|
| [29] |
D. K. Soother, J. Daudpoto, A brief review of condition monitoring techniques for the induction motor, Trans. Can. Soc. Mech. Eng., 43 (2019), 499–508. https://doi.org/10.1139/tcsme-2018-0234 doi: 10.1139/tcsme-2018-0234
|
| [30] |
A. Mehta, A. Choudhary, D. Goyal, B. S. Pabla, Infrared thermography based fault diagnosis and prognosis for rotating machines, J. Univ. Shanghai Sci. Technol., 23 (2021), 22–29. https://doi.org/10.1155/2021/9947300 doi: 10.1155/2021/9947300
|
| [31] |
S. Kavitha, N. S. Bhuvaneswari, R. Senthilkumar, N. R. Shanker, Magnetoresistance sensor-based rotor fault detection in induction motor using non-decimated wavelet and streaming data, Automatika, 63 (2022), 525–541. https://doi.org/10.1080/00051144.2022.2052533 doi: 10.1080/00051144.2022.2052533
|
| [32] | A. Ebrahimi, H. Ahmad, R. Roshanfekr, Stator winding short circuit fault detection in three-phase Induction Motors using combination type-2 Fuzzy logic and Support Vector Machine classifier optimized by Fractional-order Chaotic Particle Swarm optimization algorithm, Comput. Intell. Electr. Eng., 12 (2021), 37–48. |
| [33] |
A. Chouhan, P. Gangsar, R. Porwal, C. K. Mechefske, Artificial neural network–based fault diagnosis for induction motors under similar, interpolated and extrapolated operating conditions, Noise Vibr. Worldwide, 52 (2021), 323–333. https://doi.org/10.1177/09574565211030709 doi: 10.1177/09574565211030709
|
| [34] |
C. G. Dias, C. M. de Sousa, A neuro-fuzzy approach for locating broken rotor bars in induction motors at very low slip, J. Control Autom. Electr. Syst., 29 (2018), 489–499. https://doi.org/10.1007/s40313-018-0388-5 doi: 10.1007/s40313-018-0388-5
|
| [35] | D. Bouneb, T. Bahi, H. MERABET, Vibration for detection and diagnosis bearing faults using adaptive neurofuzzy inference system, J. Electr. Syst., 14 (2018), 95–104. |
| [36] |
Z. Zhu, Y. Lei, G. Qi, Y. Chai, N. Mazur, Y. An, et al., A review of the application of deep learning in intelligent fault diagnosis of rotating machinery, Measurement, 206 (2022), 112346. https://doi.org/10.1016/j.measurement.2022.112346 doi: 10.1016/j.measurement.2022.112346
|
| [37] |
X. Huang, G. Qi, N. Mazur, Y. Chai, Deep residual networks-based intelligent fault diagnosis method of planetary gearboxes in cloud environments, Simul. Modell. Pract. Theory, 116 (2022), 102469. https://doi.org/10.1016/j.simpat.2021.102469 doi: 10.1016/j.simpat.2021.102469
|
| [38] |
G. Qi, Z. Zhu, K. Erqinhu, Y. Chen, Y. Chai, J. Sun, Fault-diagnosis for reciprocating compressors using big data and machine learning, Simul. Modell. Pract. Theory, 80 (2018), 104–127. https://doi.org/10.1016/j.simpat.2017.10.005 doi: 10.1016/j.simpat.2017.10.005
|
| [39] |
X. Shen, G. Shi, H. Ren, W. Zhang, Biomimetic vision for zoom object detection based on improved vertical grid number YOLO algorithm, Front. Bioeng. Biotechnol., 10 (2022), 905583. https://doi.org/10.3389/fbioe.2022.905583 doi: 10.3389/fbioe.2022.905583
|
| [40] |
X. Zhang, T. Feng, Q. Niu, X. Deng, A novel swarm optimization algorithm based on a mixed-distribution model, Appl. Sci., 8 (2018), 632. https://doi.org/10.3390/app8040632 doi: 10.3390/app8040632
|
| [41] |
A. Glowacz, Thermographic fault diagnosis of shaft of BLDC motor, Sensors, 22 (2022), 8537. https://doi.org/10.3390/s22218537 doi: 10.3390/s22218537
|
| [42] |
A. Głowacz, W. Głowacz, Z. Głowacz, Recognition of armature current of DC generator depending on rotor speed using FFT, MSAF-1 and LDA, Ekspl. Niezawodność, 17 (2015), 64–69. https://doi.org/10.17531/ein.2015.1.9 doi: 10.17531/ein.2015.1.9
|
| [43] |
A. Glowacz, Fault diagnostics of acoustic signals of loaded synchronous motor using SMOFS-25-EXPANDED and selected classifiers, Tehnički vjesnik, 23 (2016), 1365–1372. https://doi.org/10.17559/TV-20150328135652 doi: 10.17559/TV-20150328135652
|
| [44] |
O. AlShorman, F. Alkahatni, M. Masadeh, M. Irfan, A. Glowacz, F. Althobiani, et al., Sounds and acoustic emission-based early fault diagnosis of induction motor: A review study, Adv. Mech. Eng., 13 (2021). https://doi.org/10.1177/1687814021996915 doi: 10.1177/1687814021996915
|
| [45] |
A. Glowacz, R. Tadeusiewicz, S. Legutko, W. Caesarendra, M. Irfan, H. Liu, et al., Fault diagnosis of angle grinders and electric impact drills using acoustic signals, Appl. Acoust., 179 (2021), 108070. https://doi.org/10.1016/j.apacoust.2021.108070 doi: 10.1016/j.apacoust.2021.108070
|








Figures(9)

Tangsheng Zhang, Hongying Zhi. A fuzzy set theory-based fast fault diagnosis approach for rotators of induction motors[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 9268-9287. doi: 10.3934/mbe.2023406







 DownLoad:
DownLoad: