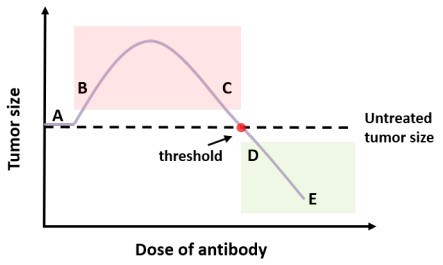
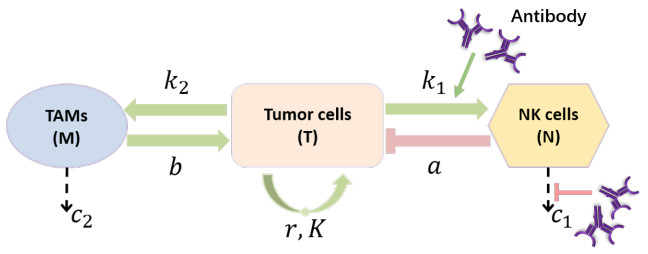
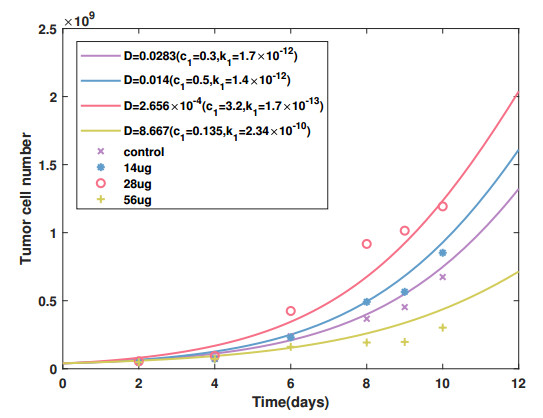
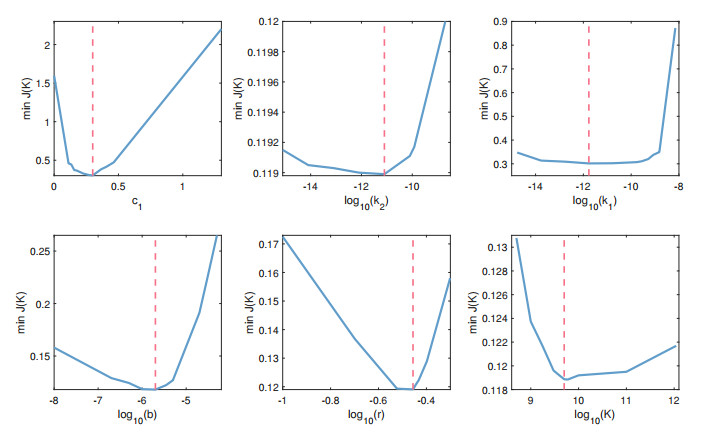
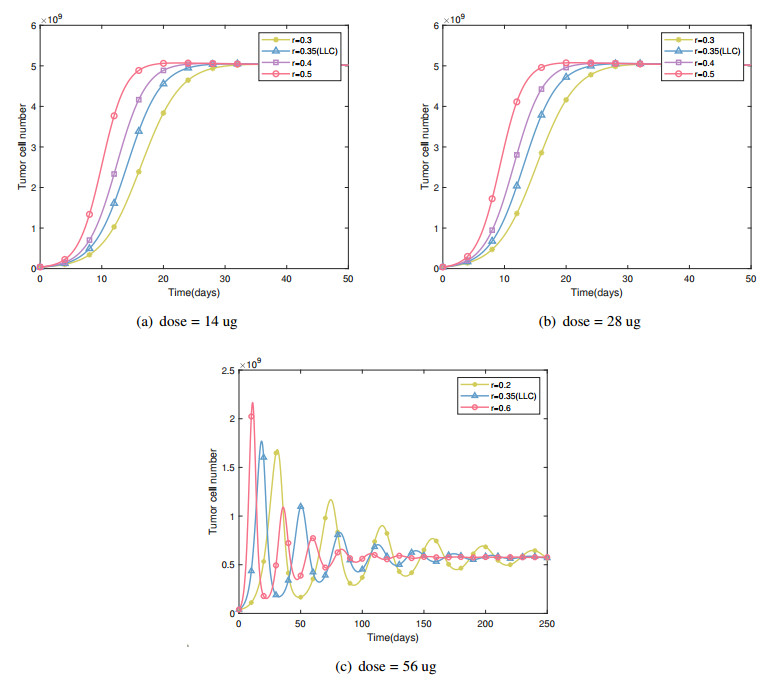
Cancer is a serious threat to human health and life. Using anti-tumor drugs is one of the important ways for treating cancer. A large number of experiments have shown that the hormesis appeared in the dose-response relationship of various anti-tumor drugs. Modeling this phenomenon will contribute to finding the appropriate dose. However, few studies have used dynamical models to quantitatively explore the hormesis phenomenon in anti-tumor drug dose-response. In this study, we present a mathematical model and dynamical analysis to quantify hormesis of anti-tumor drugs and reveal the critical threshold of antibody dose. Firstly, a dynamical model is established to describe the interactions among tumor cells, natural killer cells and M2-polarized macrophages. Model parameters are fitted through the published experimental data. Secondly, the positivity of solution and bounded invariant set are given. The stability of equilibrium points is proved. Thirdly, through bifurcation analysis and numerical simulations, the hormesis phenomenon of low dose antibody promoting tumor growth and high dose antibody inhibiting tumor growth is revealed. Furthermore, we fit out the quantitative relationship of the dose-response of antibodies. Finally, the critical threshold point of antibody dose changing from promoting tumor growth to inhibiting tumor growth is obtained. These results can provide suggestions for the selection of appropriate drug dosage in the clinical treatment of cancer.
Citation: Yuyang Xiao, Juan Shen, Xiufen Zou. Mathematical modeling and dynamical analysis of anti-tumor drug dose-response[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 4120-4144. doi: 10.3934/mbe.2022190
Cancer is a serious threat to human health and life. Using anti-tumor drugs is one of the important ways for treating cancer. A large number of experiments have shown that the hormesis appeared in the dose-response relationship of various anti-tumor drugs. Modeling this phenomenon will contribute to finding the appropriate dose. However, few studies have used dynamical models to quantitatively explore the hormesis phenomenon in anti-tumor drug dose-response. In this study, we present a mathematical model and dynamical analysis to quantify hormesis of anti-tumor drugs and reveal the critical threshold of antibody dose. Firstly, a dynamical model is established to describe the interactions among tumor cells, natural killer cells and M2-polarized macrophages. Model parameters are fitted through the published experimental data. Secondly, the positivity of solution and bounded invariant set are given. The stability of equilibrium points is proved. Thirdly, through bifurcation analysis and numerical simulations, the hormesis phenomenon of low dose antibody promoting tumor growth and high dose antibody inhibiting tumor growth is revealed. Furthermore, we fit out the quantitative relationship of the dose-response of antibodies. Finally, the critical threshold point of antibody dose changing from promoting tumor growth to inhibiting tumor growth is obtained. These results can provide suggestions for the selection of appropriate drug dosage in the clinical treatment of cancer.

| [1] |
E. J. Calabrese, Cancer biology and hormesis: Human tumor cell lines commonly display hormetic (biphasic) dose responses, Crit. Rev. Toxicol., 35 (2005), 463–582. https://doi.org/10.1080/10408440591034502 doi: 10.1080/10408440591034502

|
| [2] |
E. J. Calabrese, Hormesis: why it is important to toxicology and toxicologists, Environ. Toxicol. Chem., 27 (2008), 1451–1474. https://doi.org/10.1897/07-541.1 doi: 10.1897/07-541.1
|
| [3] | M. A. Nascarella, E. J. Stanek, G. R. Hoffmann, E. J. Calabrese, Quantification of hormesis in anticancer-agent dose-responses, Dose-Response, 7 (2009), dose–response. https://doi.org/10.2203/dose-response.08-025.Nascarella |
| [4] |
O. M. Pearce, H. Läubli, J. Bui, A. Varki, Hormesis in cancer immunology, OncoImmunology, 3 (2014), e29312. https://doi.org/10.4161/onci.29312 doi: 10.4161/onci.29312
|
| [5] |
O. M. T. Pearce, H. Laubli, A. Verhagen, P. Secrest, J. Zhang, N. M. Varki, et al., Inverse hormesis of cancer growth mediated by narrow ranges of tumor-directed antibodies, Proc. Natl. Acad. Sci. USA, 111 (2014), 5998–6003. https://doi.org/10.1073/pnas.1209067111 doi: 10.1073/pnas.1209067111
|
| [6] | T. Yoshimasu, T. Ohsahi, S. Oura, Y. Kokawa, M. Kawago, Y. Hirai, et al., A theoretical model for the hormetic dose-response curve for anticancer agents, Anticancer Res., 35 (2015), 5851–5855. |
| [7] |
Q. Li, Y. Xiao, Bifurcation analyses and hormetic effects of a discrete-time tumor model, Appl. Math. Comput., 363 (2019), 124618. https://doi.org/10.1016/j.amc.2019.124618 doi: 10.1016/j.amc.2019.124618
|
| [8] |
O. M. T. Pearce, H. Läubli, Sialic acids in cancer biology and immunity, Glycobiology, 26 (2015), 111–128. https://doi.org/10.1093/glycob/cwv097 doi: 10.1093/glycob/cwv097
|
| [9] |
M. Hedlund, V. Padler-Karavani, N. M. Varki, A. Varki, Evidence for a human-specific mechanism for diet and antibody-mediated inflammation in carcinoma progression, Proc. Natl. Acad. Sci. USA, 105 (2008), 18936–18941. https://doi.org/10.1073/pnas.0803943105 doi: 10.1073/pnas.0803943105
|
| [10] |
V. Padler-Karavani, N. Hurtado-Ziola, M. Pu, H. Yu, S. Huang, S. Muthana, et al., Human xeno-autoantibodies against a non-human sialic acid serve as novel serum biomarkers and immunotherapeutics in cancer, Cancer Res., 71 (2011), 3352–3363. https://doi.org/10.1158/0008-5472.CAN-10-4102 doi: 10.1158/0008-5472.CAN-10-4102
|
| [11] |
M. J. Smyth, Y. Hayakawa, K. Takeda, H. Yagita, New aspects of natural-killer-cell surveillance and therapy of cancer, Nat. Rev. Cancer, 2 (2002), 850–861. https://doi.org/10.1038/nrc928 doi: 10.1038/nrc928
|
| [12] |
M. Kirkilionis, S. Walcher, On comparison systems for ordinary differential equations, J. Math. Anal. Appl., 299 (2004), 157–173. https://doi.org/10.1016/j.jmaa.2004.06.025 doi: 10.1016/j.jmaa.2004.06.025
|
| [13] | H. Khalil, Nonlinear Systems, Prentice Hall, 2002. |
| [14] | Q. Rahman, Analytic Theory of Polynomials, Clarendon Press Oxford University Press, Oxford New York, 2002. |
| [15] |
U. Moran, R. Phillips, R. Milo, Snapshot: Key numbers in biology, Cell, 141 (2010), 1262–1262.e1. https://doi.org/10.1016/j.cell.2010.06.019 doi: 10.1016/j.cell.2010.06.019
|
| [16] |
A. M. Lutz, J. K. Willmann, F. V. Cochran, P. Ray, S. S. Gambhir, Cancer Screening: A mathematical model relating secreted blood biomarker levels to tumor sizes, PLoS Med., 5 (2008), e170. https://doi.org/10.1371/journal.pmed.0050170 doi: 10.1371/journal.pmed.0050170
|
| [17] | E. Russell, K. James, Particle swarm optimization, in Proceedings of the IEEE international conference on neural networks, 4 (1995), 1942–1948, https://doi.org/10.1109/ICNN.1995.488968 |
| [18] |
L. G. de Pillis, A. E. Radunskaya, C. L. Wiseman, A validated mathematical model of cell-mediated immune response to tumor growth, Cancer Res., 65 (2005), 7950–7958. https://doi.org/10.1158/0008-5472.CAN-05-0564 doi: 10.1158/0008-5472.CAN-05-0564
|
| [19] |
B. Mukhopadhyay, R. Bhattacharyya, Temporal and spatiotemporal variations in a mathematical model of macrophage-tumor interaction, Nonlinear Anal. Hybrid Syst., 2 (2008), 819–831. https://doi.org/10.1016/j.nahs.2007.11.011 doi: 10.1016/j.nahs.2007.11.011
|
| [20] |
C. Kreutz, A. Raue, J. Timmer, Likelihood based observability analysis and confidence intervals for predictions of dynamic models, BMC Syst. Biol., 6 (2012), 120. https://doi.org/10.1186/1752-0509-6-120 doi: 10.1186/1752-0509-6-120
|
| [21] |
A. Raue, C. Kreutz, T. Maiwald, J. Bachmann, M. Schilling, U. Klingmüller, et al., Structural and practical identifiability analysis of partially observed dynamical models by exploiting the profile likelihood, Bioinformatics, 25 (2009), 1923–1929. https://doi.org/10.1093/bioinformatics/btp358 doi: 10.1093/bioinformatics/btp358
|
| [22] |
I. Borisov, E. Metelkin, Confidence intervals by constrained optimization—An algorithm and software package for practical identifiability analysis in systems biology, PLOS Comput. Biol., 16 (2020), e1008495. https://doi.org/10.1371/journal.pcbi.1008495 doi: 10.1371/journal.pcbi.1008495
|
| [23] |
X. Zou, X. Xiang, Y. Chen, T. Peng, X. Luo, Z. Pan, Understanding inhibition of viral proteins on type i IFN signaling pathways with modeling and optimization, J. Theor. Biol., 265 (2010), 691–703. https://doi.org/10.1016/j.jtbi.2010.05.001 doi: 10.1016/j.jtbi.2010.05.001
|
| [24] |
S. Y. Shin, O. Rath, S. M. Choo, F. Fee, B. McFerran, W. Kolch, et al., Positive- and negative-feedback regulations coordinate the dynamic behavior of the ras-raf-MEK-ERK signal transduction pathway, J. Cell Sci., 122 (2009), 425–435. https://doi.org/10.1242/jcs.036319 doi: 10.1242/jcs.036319
|
| [25] |
P. R. Crocker, J. C. Paulson, A. Varki, Siglecs and their roles in the immune system, Nat. Rev. Immunol., 7 (2007), 255–266. https://doi.org/10.1038/nri2056 doi: 10.1038/nri2056
|
Figures(13) / Tables(6)

Yuyang Xiao, Juan Shen, Xiufen Zou. Mathematical modeling and dynamical analysis of anti-tumor drug dose-response[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 4120-4144. doi: 10.3934/mbe.2022190







 DownLoad:
DownLoad: