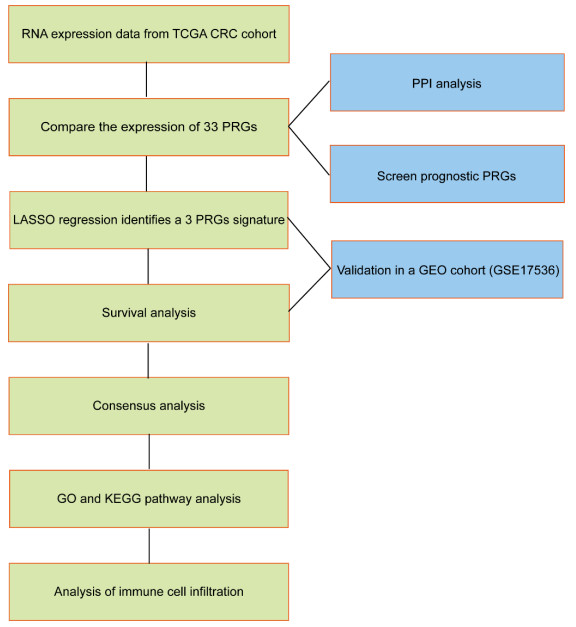
Colorectal cancer (CRC), one of the most common malignancies worldwide, leads to abundant cancer-related mortalities annually. Pyroptosis, a new kind of programmed cell death, plays a critical role in immune response and tumor progression. Our study aimed to identify a prognostic signature for CRC based on pyroptosis-related genes (PRGs). The difference in PRGs between CRC tissues and normal tissues deposited in the TCGA database was calculated by "limma" R package. The tumor microenvironment (TME) of CRC cases was accessed by the ESTIMATE algorithm. The prognostic PRGs were identified using Cox regression analysis. A least absolute shrinkage and selector operation (LASSO) algorithm was used to calculate the risk scores and construct a clinical predictive model of CRC. Gene Set Enrichment Analysis (GSEA) was performed for understanding the function annotation of the signature in the tumor microenvironment. We found that most PRGs were significantly dysregulated in CRC. Through the LASSO method, three key PRGs were selected to calculate the risk scores and construct the prognostic model for CRC. The risk score was an independent indicator of patient's prognosis. In addition, we classified the CRC patients into two clusters based on risk scores and discovered that CRC patients in cluster 2 underwent worse overall survival and owned higher expression levels of immune checkpoint genes in tumor tissues. In conclusion, our study identified a PRG-related prognostic signature for CRC, according to which we classified the CRC patients into two clusters with distinct prognosis and immunotherapy potential.
Citation: Chen Zheng, Zhaobang Tan. A novel identified pyroptosis-related prognostic signature of colorectal cancer[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8783-8796. doi: 10.3934/mbe.2021433
Colorectal cancer (CRC), one of the most common malignancies worldwide, leads to abundant cancer-related mortalities annually. Pyroptosis, a new kind of programmed cell death, plays a critical role in immune response and tumor progression. Our study aimed to identify a prognostic signature for CRC based on pyroptosis-related genes (PRGs). The difference in PRGs between CRC tissues and normal tissues deposited in the TCGA database was calculated by "limma" R package. The tumor microenvironment (TME) of CRC cases was accessed by the ESTIMATE algorithm. The prognostic PRGs were identified using Cox regression analysis. A least absolute shrinkage and selector operation (LASSO) algorithm was used to calculate the risk scores and construct a clinical predictive model of CRC. Gene Set Enrichment Analysis (GSEA) was performed for understanding the function annotation of the signature in the tumor microenvironment. We found that most PRGs were significantly dysregulated in CRC. Through the LASSO method, three key PRGs were selected to calculate the risk scores and construct the prognostic model for CRC. The risk score was an independent indicator of patient's prognosis. In addition, we classified the CRC patients into two clusters based on risk scores and discovered that CRC patients in cluster 2 underwent worse overall survival and owned higher expression levels of immune checkpoint genes in tumor tissues. In conclusion, our study identified a PRG-related prognostic signature for CRC, according to which we classified the CRC patients into two clusters with distinct prognosis and immunotherapy potential.

| [1] |
R. L. Siegel, K. D. Miller, A. Goding Sauer, S. A. Fedewa, L. F. Butterly, J. C. Anderson, et al., Colorectal cancer statistics, 2020, CA Cancer J. Clin, 70 (2020), 145-164. doi: 10.3322/caac.21601

|
| [2] |
S. Dhillon, Regorafenib: A review in metastatic colorectal cancer, Drugs, 78 (2018), 1133-1144. doi: 10.1007/s40265-018-0938-y
|
| [3] | Y. Fang, S. Tian, Y. Pan, W. Li, Q. Wang, Y. Tang, et al., Pyroptosis: A new frontier in cancer, Biomed. Pharm., 121 (2020), 109595. |
| [4] | R. Tang, J. Xu, B. Zhang, J. Liu, C. Liang, J. Hua, et al., Ferroptosis, necroptosis, and pyroptosis in anticancer immunity, J. Hematol. Oncol, 13 (2020), 110. |
| [5] |
L. S. Wu, Y. Liu, X. W. Wang, B. Xu, Y. L. Lin, Y. Song, et al., LPS enhances the chemosensitivity of oxaliplatin in HT29 cells via GSDMD-mediated pyroptosis, Cancer Manag. Res., 12 (2020), 10397-10409. doi: 10.2147/CMAR.S244374
|
| [6] |
J. Guo, J. Zheng, M. Mu, Z. Chen, Z. Xu, C. Zhao, et al., GW4064 enhances the chemosensitivity of colorectal cancer to oxaliplatin by inducing pyroptosis, Biochem. Biophys. Res. Commun., 548 (2021), 60-66. doi: 10.1016/j.bbrc.2021.02.043
|
| [7] | Z. Tang, L. Ji, M. Han, J. Xie, F. Zhong, X. Zhang, et al., Pyroptosis is involved in the inhibitory effect of FL118 on growth and metastasis in colorectal cancer, Life Sci., 257 (2020), 118065. |
| [8] |
Z. Zhang, Y. Zhang, S. Xia, Q. Kong, S. Li, X. Liu, et al., Gasdermin E suppresses tumour growth by activating anti-tumour immunity, Nature, 579 (2020), 415-420. doi: 10.1038/s41586-020-2071-9
|
| [9] |
D. A. Erkes, W. Cai, I. M. Sanchez, T. J. Purwin, C. Rogers, C. O. Field, et al., Mutant BRAF and MEK inhibitors regulate the tumor immune microenvironment via pyroptosis, Cancer Discov., 10 (2020), 254-269. doi: 10.1158/2159-8290.CD-19-0672
|
| [10] |
M. S. Chen, Y. H. Lo, X. Chen, C. S. Williams, J. M. Donnelly, Z. K. Criss, et al., Growth factor-independent 1 is a tumor suppressor gene in colorectal cancer, Mol. Cancer Res., 17 (2019), 697-708. doi: 10.1158/1541-7786.MCR-18-0666
|
| [11] | W. Lin, Y. Chen, B. Wu, Y. Chen, Z. Li, Identification of the pyroptosis-related prognostic gene signature and the associated regulation axis in lung adenocarcinoma, Cell Death Discov., 7 (2021), 161. |
| [12] |
H. Kambara, F. Liu, X. Zhang, P. Liu, B. Bajrami, Y. Teng, et al., Gasdermin D exerts anti-inflammatory effects by promoting neutrophil death, Cell Rep., 22 (2018), 2924-2936. doi: 10.1016/j.celrep.2018.02.067
|
| [13] |
B. Zech, R. Köhl, A. von Knethen, B. Brüne, Nitric oxide donors inhibit formation of the Apaf-1/caspase-9 apoptosome and activation of caspases, Biochem. J., 371 (2003), 1055-1064. doi: 10.1042/bj20021720
|
| [14] |
A. Liberzon, C. Birger, H. Thorvaldsdóttir, M. Ghandi, J. P. Mesirov, P. Tamayo, The Molecular Signatures Database (MSigDB) hallmark gene set collection, Cell Syst., 1 (2015), 417-425. doi: 10.1016/j.cels.2015.12.004
|
| [15] |
W. Huang da, B. T. Sherman, R. A. Lempicki, Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources, Nat. Protoc., 4 (2009), 44-57. doi: 10.1038/nprot.2008.211
|
| [16] |
G. Bindea, B. Mlecnik, M. Tosolini, A. Kirilovsky, M. Waldner, A. C. Obenauf, et al., Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer, Immunity, 39 (2013), 782-795. doi: 10.1016/j.immuni.2013.10.003
|
| [17] | M. S. D'Arcy, Cell death: A review of the major forms of apoptosis, necrosis and autophagy, Cell Biol. Int., 43 (2019), 582-592. |
| [18] | X. Xia, X. Wang, Z. Cheng, W. Qin, L. Lei, J. Jiang, et al., The role of pyroptosis in cancer: Pro-cancer or pro-"host"?, Cell Death Dis, 10 (2019), 650. |
| [19] |
N. Saeki, T. Usui, K. Aoyagi, D. H. Kim, M. Sato, T. Mabuchi, et al., Distinctive expression and function of four GSDM family genes (GSDMA-D) in normal and malignant upper gastrointestinal epithelium, Genes Chromosomes Cancer, 48 (2009), 261-271. doi: 10.1002/gcc.20636
|
| [20] | M. Miguchi, T. Hinoi, M. Shimomura, T. Adachi, Y. Saito, H. Niitsu, et al., Gasdermin C is upregulated by inactivation of transforming growth factor β receptor type Ⅱ in the presence of mutated Apc, promoting colorectal cancer proliferation, PLoS One, 11 (2016), e0166422. |
| [21] | J. Gao, X. Qiu, G. Xi, H. Liu, F. Zhang, T. Lv, et al., Downregulation of GSDMD attenuates tumor proliferation via the intrinsic mitochondrial apoptotic pathway and inhibition of EGFR/Akt signaling and predicts a good prognosis in non-small cell lung cancer, Oncol. Rep., 40 (2018), 1971-1984. |
| [22] |
I. C. Allen, E. M. TeKippe, R. M. Woodford, J. M. Uronis, E. K. Holl, A. B. Rogers, et al., The NLRP3 inflammasome functions as a negative regulator of tumorigenesis during colitis-associated cancer, J. Exp. Med., 207 (2010), 1045-1056. doi: 10.1084/jem.20100050
|
| [23] | C. Chen, B. Wang, J. Sun, H. Na, Z. Chen, Z. Zhu, et al., DAC can restore expression of NALP1 to suppress tumor growth in colon cancer, Cell Death Dis., 6 (2015), e1602. |
| [24] |
S. Dihlmann, S. Tao, F. Echterdiek, E. Herpel, L. Jansen, J. Chang-Claude, et al., Lack of Absent in Melanoma 2 (AIM2) expression in tumor cells is closely associated with poor survival in colorectal cancer patients, Int. J. Cancer, 135 (2014), 2387-2396. doi: 10.1002/ijc.28891
|
| [25] |
J. Ibrahim, K. Op de Beeck, E. Fransen, L. Croes, M. Beyens, A. Suls, et al., Methylation analysis of Gasdermin E shows great promise as a biomarker for colorectal cancer, Cancer Med., 8 (2019), 2133-2145. doi: 10.1002/cam4.2103
|
| [26] | W. Shao, Z. Yang, Y. Fu, L. Zheng, F. Liu, L. Chai, et al., The Pyroptosis-related signature predicts prognosis and indicates immune microenvironment infiltration in gastric cancer, Front. Cell Dev. Biol., 9 (2021), 676485. |
| [27] | Y. Ye, Q. Dai, H. Qi, A novel defined pyroptosis-related gene signature for predicting the prognosis of ovarian cancer, Cell Death Discov., 7 (2021), 71. |
| [28] | R. Schwarzer, L. Laurien, M. Pasparakis, New insights into the regulation of apoptosis, necroptosis, and pyroptosis by receptor interacting protein kinase 1 and caspase-8, Curr. Opin. Cell. Biol., 63 (2020), 186-193. |
| [29] |
Q. Wang, Y. Wang, J. Ding, C. Wang, X. Zhou, W. Gao, et al., A bioorthogonal system reveals antitumour immune function of pyroptosis, Nature, 579 (2020), 421-426. doi: 10.1038/s41586-020-2079-1
|
| [30] |
A. Rizzo, M. Di Giovangiulio, C. Stolfi, E. Franzè, H. J. Fehling, R. Carsetti, et al., RORγt-expressing tregs drive the growth of colitis-associated colorectal cancer by controlling IL6 in dendritic cells, Cancer Immunol. Res., 6 (2018), 1082-1092. doi: 10.1158/2326-6066.CIR-17-0698
|
| [31] | S. Zhao, Y. Mi, B. Guan, B. Zheng, P. Wei, Y. Gu, et al., Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer, J. Hematol. Oncol., 13 (2020), 156. |
| [32] |
B. Li, H. L. Chan, P. Chen, Immune checkpoint inhibitors: Basics and challenges, Curr. Med. Chem., 26 (2019), 3009-3025. doi: 10.2174/0929867324666170804143706
|
| [33] |
Y. Y. Gu, M. H. Chen, B. H. May, X. Z. Liao, J. H. Liu, L. T. Tao, et al., Matrine induces apoptosis in multiple colorectal cancer cell lines in vitro and inhibits tumour growth with minimum side effects in vivo via Bcl-2 and caspase-3, Phytomedicine, 51 (2018), 214-225. doi: 10.1016/j.phymed.2018.10.004
|
| [34] | Q. Han, Y. Ma, H. Wang, Y. Dai, C. Chen, Y. Liu, et al., Resibufogenin suppresses colorectal cancer growth and metastasis through RIP3-mediated necroptosis, J. Transl. Med., 16 (2018), 201. |
| [35] | S. Christgen, M. Zheng, S. Kesavardhana, R. Karki, R. K. S. Malireddi, B. Banoth, et al., Identification of the PANoptosome: A molecular platform triggering pyroptosis, apoptosis, and necroptosis (PANoptosis), Front. Cell Infect. Microbiol., 10 (2020), 237. |
| [36] |
I. Jorgensen, M. Rayamajhi, E. A. Miao, Programmed cell death as a defence against infection, Nat. Rev. Immunol., 17 (2017), 151-164. doi: 10.1038/nri.2016.147
|





Figures(6) / Tables(1)

Chen Zheng, Zhaobang Tan. A novel identified pyroptosis-related prognostic signature of colorectal cancer[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8783-8796. doi: 10.3934/mbe.2021433







 DownLoad:
DownLoad: