Citation: Rachael C. Adams, Behnam Rashidieh. Can computers conceive the complexity of cancer to cure it? Using artificial intelligence technology in cancer modelling and drug discovery[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6515-6530. doi: 10.3934/mbe.2020340

| [1] | R. Mahumad, K. Alam, J. Dunn, J. Gow, Emerging cancer incidence, mortality, hospitalisation and associated burden among Australian cancer patients, 1982-2014: An incidence-based approach in terms of trends, determinants and inequality, BMJ Open, 5 (2019). |
| [2] | M. Breitenbach, J. Hoffmann, Editorial: Cancer models, Front. Oncol., 8 (2018), 401-401. |
| [3] | L. Ogilvie, A. Kovachev, C. Wierling, B. Lange, H. Lehrach, Models of models: A translational Route for cancer treatment and drug development, Front. Oncol., 7 (2017). |
| [4] | K. Mak, M. Pichika, Artificial intelligence in drug development: Present status and future prospects, Drug Discovery Today, 24 (2019), 773-780. |
| [5] | J. Vamathevan, D. Clark, P. Czodrowski, I. Dunham, E. Ferran, G. Lee, et al., Applications of machine learning in drug discovery and development, Nat. Rev. Drug Discovery, 18 (2018), 463-477. |
| [6] | O. Wolkenhauer, Why model? Front. Phys., 5 (2014). |
| [7] | A. Levine, C. Schlosser, J. Grewal., R. Coope, S. Jones, S. Yip, Rise of the machines: Advances in deep learning for cancer diagnosis, Trends Cancer, 5 (2019), 157-169. |
| [8] | K. Vougas, T. Sakellaropolous, A. Kotsina, G. R. P. Foukas, A. Ntargaras, F. Koinis, et al., Machine learning and data mining frameworks for predicting drug response in cancer: An overview and a novel in silico screening process based on association rule mining. Pharmacol. Ther., 203 (2019), 107395. |
| [9] | G. V. Sherbet, W. L. Woo, S. Dlay, Application of artificial intelligence-based technology in cancer management: A commentary on the deployment of artificial neural networks, Anticancer Res., 38 (2018), 6607-6613. |
| [10] | R. M. Thomas, T. Van Dyke, G. Merlino, C. P. Day, Concepts in cancer modeling: A brief history, Cancer Res., 76 (2016), 5921-5925. |
| [11] | P. Kumari, A. Nath, R. Chaube, Identification of human drug targets using machine-learning algorithms, Comput. Biol. Med., 56 (2014), 175-181. |
| [12] | J. Metzcar, Y. Wang, R. Heiland, P. Macklin, A Review of cell-based computational modeling in cancer biology, JCO Clin. Cancer Inform., 3 (2019), 1-13. |
| [13] | D. Hanahan, R. A. Weinberg, The hallmarks of cancer, Cell, 100 (2000), 57-70. |
| [14] | A. Ghaffarizadeh, S. H. Friedman, P. Macklin, BioFVM: An efficient, parallelized diffusive transport solver for 3-D biological simulations, Bioinformatics, 32 (2016), 1256-1258. |
| [15] | A. Ghaffarizadeh, R. Heiland, S. H. Friedman, S. M. Mumethaler, P. Macklin, PhysiCell: An open source physics-based cell simulator for 3-D multicellular systems, PLOS Comput. Biol., 14 (2018), e1005991. |
| [16] | E. Bonabeau, Agent-based modeling: Methods and techniques for simulating human systems, Proc. Natl. Acad. Sci. USA, 99 (2002), 7280-7287. |
| [17] | P. Van Liedekerke, M. M. Palm, N. Jagiella, D. Drasdo, Simulating tissue mechanics with agent-based models: Concepts, perspectives and some novel results, Comput. Particle Mech., 2 (2015), 401-444. |
| [18] | R. C. Kennedy, G. E. Ropella, C. A. Hunt, A cell-centered, agent-based framework that enables flexible environment granularities, Theor. Biol. Med. Model. 13 (2016). |
| [19] | J. Poleszczuk, P. Macklin, H. Enderling, Agent-based modeling of cancer stem cell driven solid tumor growth, Methods Mol. Biol., 1516 (2016), 335-346. |
| [20] | Y. Cai, S. Xu, J. Wu, Q. Long, Coupled modelling of tumour angiogenesis, tumour growth and blood perfusion, J. Theor. Biol., 279 (2011), 90-101. |
| [21] | A. Shirinifard, J. S. Gens, B. L. Zaitlen, N. J. Poplawski, M. Swat, J. A. Glazier, 3D multi-cell simulation of tumor growth and angiogenesis, PLOS One, 4 (2009), e7190. |
| [22] | B. Chopard, R. Ouared, A. Deustch, H. Hatzikirou, D. Wolf-Gladrow, Lattice-gas cellular automaton models for biology: From fluids to cells, Acta Biotheor., 58 (2010), 329-340. |
| [23] | H. Hatzikirou, D. Basanta, M. Simon, K. Schaller, A. Deustch, 'Go or Grow': The key to the emergence of invasion in tumour progression?, Math. Med. Biol.: A J. IMA, 29 (2010), 49-65. |
| [24] | H. N. Weerasinghe, P. M. Burragem, K. Burrage, D. V. Nicolau, Mathematical models of cancer cell plasticity, J. Oncol., 2019. |
| [25] | M. S. Alber, M. A. Kiskowski, J. A. Glazier, Y. Jiang, On cellular automaton approaches to modeling biological cells, in Mathematical Systems Theory in Biology, Communications, Computation, and Finance, Springer, New York, (2003), 1-39. |
| [26] | A. Szabó, R. M. Merks, Cellular potts modeling of tumor growth, tumor invasion, and tumor evolution, Front. Oncol., 3 (2013). |
| [27] | N. Guisoni, K. I. Mazzitello, L. Diambra, Modeling active cell movement with the potts model, Front. Phys., 6 (2018). |
| [28] | E. G. Rens, L. Edelstein-Keshet, From energy to cellular forces in the cellular potts model: An algorithmic approach, PLOS Comput. Biol., 15 (2019), e1007459. |
| [29] | K. A. Rejniak, A. R. A. Anderson, Hybrid models of tumor growth, Wiley Interdiscip. Rev.: Syst. Biol. Med., 3 (2011), 115-125. |
| [30] | J. Jeon, V. Quaranta, P. T. Cummings, An off-lattice hybrid discrete-continuum model of tumor growth and invasion, Biophys. J., 98 (2010), 37-47. |
| [31] | S. Koride, A. J. Loza, S. X. Sun, Epithelial vertex models with active biochemical regulation of contractility can explain organized collective cell motility, APL Bioeng., 2 (2018). |
| [32] | S. Alt, P. Ganguly, G. Salbreux, Vertex models: from cell mechanics to tissue morphogenesis, Philos. Tran. R. Soc. London. Ser. B, Biol. Sci., 372 (2017). |
| [33] | K. R. Foster, R. Koprowski, J. D. Skufca, Machine learning, medical diagnosis, and biomedical engineering research-commentary, Biomed. Eng. Online, 13 (2014), 94. |
| [34] | A. M. Shirin, S. S. Dlay, W. L. Woo, G. V. Sherbet, Cross validation evaluation for breast cancer prediction using multilayer perceptron neural networks, Am. J. Eng. Appl. Sci., 4 (2012). |
| [35] | T. Dietterich, Overfitting and undercomputing in machine learning, ACM Comput. Surveys, 27 (1995), 326-327. |
| [36] | T. L. Whiteside, The tumor microenvironment and its role in promoting tumor growth, Oncogene, 27 (2018), 5904-5912. |
| [37] | D. Hanahan, R. A. Weinberg, Hallmarks of cancer: the next generation, Cell, 144 (2011), 646-674. |
| [38] | A. M. Newman, C. B. Steen, C. L. Liu, A. J. Gentles, A. A. Chaudhuri, F. Scherer, et al., Determining cell type abundance and expression from bulk tissues with digital cytometry. Nat. Biotech., 37 (2019), 773-782. |
| [39] | B. Chen, M. S. Khodadoust, C. L. Liu, A. M. Newman, A. A. Alizadeh, Profiling tumor infiltrating immune cells with CIBERSORT, Methods Mol. Biol., 1711 (2018), 243-259. |
| [40] | K. Menden, M. Marouf, S. Oller, A. Dalmia, D. S. Magruder, K. Kloiber, et al., Deep learning-based cell composition analysis from tissue expression profiles, Sci. Adv., 6 (20100), eaba2619. |
| [41] | X. Sun, B. Hu, Mathematical modeling and computational prediction of cancer drug resistance, Brief Bioinf., 19 (2018), 1382-1399. |
| [42] | J. Cosgrove, J. Butler, K. Alden, M. Read, V. Kumar, L. Cucurull-Sanchez, et al., Agent-based modeling in systems pharmacology, CPT: Pharmacometrics Syst. Pharmacol., 4 (2015), 615-629. |
| [43] | R. L. Dedrick, D. S. Zaharko, R. A. Bender, W. A. Bleyer, R. J. Lutz, Pharmacokinetic considerations on resistance to anticancer drugs, Cancer Chemother Rep., 59 (1975), 795-804. |
| [44] | H. B. Frieboes, M. E. Edgerton, J. P. Fruehauf, F. R. A. J. Rose, L. K. Worrall, R. A. Gatenby, et al., Prediction of drug response in breast cancer using integrative experimental/computational modeling, Cancer Res., 69 (2009, 4484-4492. |
| [45] | B. G. Birkhead, E. M. Rankin, S. Gallivan, L. Dones, R. D. Rubens, A mathematical model of the development of drug resistance to cancer chemotherapy, Eur. J. Cancer Clin. Oncol., 23 (1987), 1421-1427. |
| [46] | A. Ghaffarizadeh, S. H. Friedman, P. Macklin, BioFVM: an efficient, parallelized diffusive transport solver for 3-D biological simulations, Bioinformatics, 32 (2015), 1256-1258. |
| [47] | E. F. Juarez, R. Lau, S. H. Friedman, A. Ghaffarizadeh, E. Jonckheere, D. B. Agus, et al., Quantifying differences in cell line population dynamics using CellPD, BMC Syst. Biol., 10 (2016). |
| [48] | S. H. Friedman, A. R. A. Anderson, D. M. Bortz, A. G. Fletcher, H. B. Frieboes, A. Ghaffarizadeh, et al., MultiCellDS: a standard and a community for sharing multicellular data, preprint, bioRxiv 090696. |
| [49] | H. C. Tang, Y. C. Chen, Insight into molecular dynamics simulation of BRAF(V600E) and potent novel inhibitors for malignant melanoma, Int. J. Nanomedicine, 10 (2015), 3131-3146. |
| [50] | K. J. Mahasa, R. Ouifki, A. Eladdadi, L. de Pillis, Mathematical model of tumor-immune surveillance, J. Theor. Biol., 404 (2016), 312-330. |
| [51] | T. Jackson, A. Radunskaya, Applications of Dynamical Systems in Biology and Medicine, 2015. |
| [52] | A. Dhawan, T. A. Graham, A. G. Fletcher, A computational modeling approach for deriving biomarkers to predict cancer risk in premalignant disease, Cancer Prev. Res., 9 (2016), 283-295. |
| [53] | X. Yang, Y. Wang, R. Byrne, G. Schneider, S. Yang, Concepts of artificial intelligence for computer-assisted drug discovery, Chem. Rev., 119 (2019), 10520-10594. |
| [54] | W. Kolch, D. Fey, Personalized computational models as biomarkers, J. Pers. Med., 7 (2017), 9. |
| [55] | C. L. Fischer, A. M. Bates, E. A. Lanzel, J. M. Guthmiller, G. K. Johnson, N. K. Singh, et al., Computational models accurately predict multi-cell biomarker profiles in inflammation and cancer, Sci. Rep., 9 (2019), 10877. |
| [56] | F. J. Esteva, G. N. Hortobagyi, Prognostic molecular markers in early breast cancer, Breast Cancer Res., 6 (2004), 109-118. |
| [57] | S. Mojarad, B. Venturini, P. Fulgenzi, R. Papaleo, M. Brisigotti, F. Monti, et al., Prediction of nodal metastasis and prognosis of breast cancer by ANN-based assessment of tumour size and p53, Ki-67 and steroid receptor expression, Anticancer Res., 33 (2013), 3925-3933. |
| [58] | M. P. Menden, F. Iorio, M. Garnett, U. McDermott, C. H. Benes, P. J. Ballester, et al., Machine learning prediction of cancer cell sensitivity to drugs based on genomic and chemical properties, Plos One, 8 (2013), e61318. |
| [59] | A. Bravo, J. Pinero, N. Queralt-Rosinach, M. Rautschka, L. I. Furlong, Extraction of relations between genes and diseases from text and large-scale data analysis: implications for translational research, BMC Bioinfor., 16 (2015). |
| [60] | J. Kim, J-j. Kim, H. Lee, An analysis of disease-gene relationship from medline abstracts by DigSee, Sci. Rep., 7 (2017), 40154. |
| [61] | P. Mamoshina, M. Volosnikova, I. V. Ozerov, E. Putin, E. Skibina, F. Cortese, et al., Machine learning on human muscle transcriptomic data for biomarker discovery and tissue-specific drug target identification, Front. Genet., 9 (2018). |
| [62] | T. Zhu, S, Cao, P. C. Su, R. Patel, D. Shah, H. B. Chokshi, et al., Hit identification and optimization in virtual screening: Practical recommendations based on a critical literature analysis, J. Med. Chem., 56 (2013), 6560-6572. |
| [63] | G. Klopman, S. K. Chakravarti, H, Zhu, J. M. Ivanov, R. D. Saiakhov, ESP: A method to predict toxicity and pharmacological properties of chemicals using multiple MCASE databases, J. Chem. Infor. Comput. Sci., 44 (2004), 704-715. |
| [64] | I. W. Mak, N. Evaniew, M. Ghert, Lost in translation: animal models and clinical trials in cancer treatment, Am. J. Trans. Res., 6 (2014), 114-118. |
| [65] | B. Ramsundar, B. Liu, Z. Wu, A. Verra, M. Tudor, R. P. Sherridan, et al., Is multitask deep learning practical for pharma?, J. Chem. Infor. Model., 57 (2017), 2068-2076. |
| [66] | M. Olivecrona, T. Blashcke, O. Engvist, H. Chen, Molecular de-novo design through deep reinforcement learning, J. Cheminfor., 9 (2017). |
| [67] | T. Luechtefeld, D. Marsh, C. Rowlands, T. Hartung, Machine learning of toxicological big data enables read-across structure activity relationships (RASAR) outperforming animal test reproducibility, Toxicol. Sci., 165 (2018), 198-212. |
| [68] | J. L. Perez-Gracia, M. F. Sanmamed, A. Bosch, A. Patino-Garcia, K. A. Schalper, V. Segura, et al., Strategies to design clinical studies to identify predictive biomarkers in cancer research, Cancer Treat. Rev., 53 (2017), 79-97. |
| [69] | E. E. Bain, L. SHafner, D. P. Walling, A. A. Othman, C. C. Stein, J. Hinkle, et al., Use of a novel artificial intelligence platform on mobile devices to assess dosing compliance in a Phase 2 clinical trial in subjects with schizophrenia, JMIR mHealth uHealth, 5 (2017), e18. |
| [70] | A. H. Beck, A. R. Sangoi, S. Leung, R. K. Marinelli, T. O. Nielsen, M. J. van de Vijver, et al., Systematic analysis of breast cancer morphology uncovers stromal features associated with survival, Sci. Trans. Med., 3 (2011). |
| [71] | N. L. Mani, K. A. Schalper, C. Hatzis, O. Saglam, F. Tavassoli, M. Butler, et al., Quantitative assessment of the spatial heterogeneity of tumor-infiltrating lymphocytes in breast cancer, Breast Cancer Res., 18 (2016). |
| [72] | F. Lake, Artificial intelligence in drug discovery: what is new, and what is next?, Future Drug Discovery, 1 (2019). |
| [73] | K. L. Fetah, B. J. DiPArdo, E. M. Kongadzem, J. S. Tomlinson, A. Elzaghied, M. Elmusrati, et al., Cancer modeling-on-a-chip with future artificial intelligence integration, Small, 15 (2019). |
| [74] | V. Assadollahi, B. Rashidieh, M. Alasvand, A. Abdolahi, J. A. Lopez, Interaction and molecular dynamics simulation study of Osimertinib (AstraZeneca 9291) anticancer drug with the EGFR kinase domain in native protein and mutated L844V and C797S, J. Cell. Biochem., 120 (2019), 13046-13055. |
| [75] | S. Ghafari, M. Komeilian, M. Hashemi, S. Oushani, G. Rigi, B. Rashidieh, et al., Molecular docking based screening of Listeriolysin-O for improved inhibitors, Bioinformation, 13 (2017), 160-163. |
| [76] | V. Assadollahi, B. Rashidieh, Molecular dynamics simulation of EFGR L844V mutant sensitive to AZD9291 in non-small cell lung cancer, J. Thorac. Oncol., 12 (2017), 1210. |
| [77] | M. M. Ranbar, V. Assadolahi, M. Yazdani, D. Nikaein, B. Rashidieh, Virtual dual inhibition of COX-2/5-LOX enzymes based on binding properties of alpha-amyrins, the anti-inflammatory compound as a promising anti-cancer drug, EXCLI J., 15 (2016), 238-245. |
| [78] | B. Rashidieh, M. Valizadeh, V. Assadollahi, M. M. Ranjbar, Molecular dynamics simulation on the low sensitivity of mutants of NEDD-8 activating enzyme for MLN4924 inhibitor as a cancer drug, Am. J. Cancer Res., 5 (2015), 3400-3406. |
| [79] | B. Rashidieh, Z. Madani. M. K. Azam, S. K. Maklavani, N. R. Akbari, S. Tavakoli, et al., Molecular docking based virtual screening of compounds for inhibiting sortase A in L. monocytogenes, Bioinformation, 11 (2015), 501-505. |
| [80] | B. Rashidieh, S. Etemadiafshar, G. Memari, M. Mirzaeichegeni, S. Yazdi, F. Farsimadan, et al., A molecular modeling based screening for potential inhibitors to alpha hemolysin from Staphylococcus aureus, Bioinformation, 11 (2015), 373-377. |
| [81] | J. Ozik, N. Collier, R. Heiland, G. An, P. Macklin, Learning-accelerated discovery of immune-tumour interactions, Mol. Syst. Des. Eng., 4 (2019), 747-760. |
| [82] | J. Ozik, N. Collier, J. M. Wozniak, C. Macal, C. Cockrell, S. F. Friedman, et al., High-throughput cancer hypothesis testing with an integrated PhysiCell-EMEWS workflow, BMC Bioinfor., 19 (2018). |
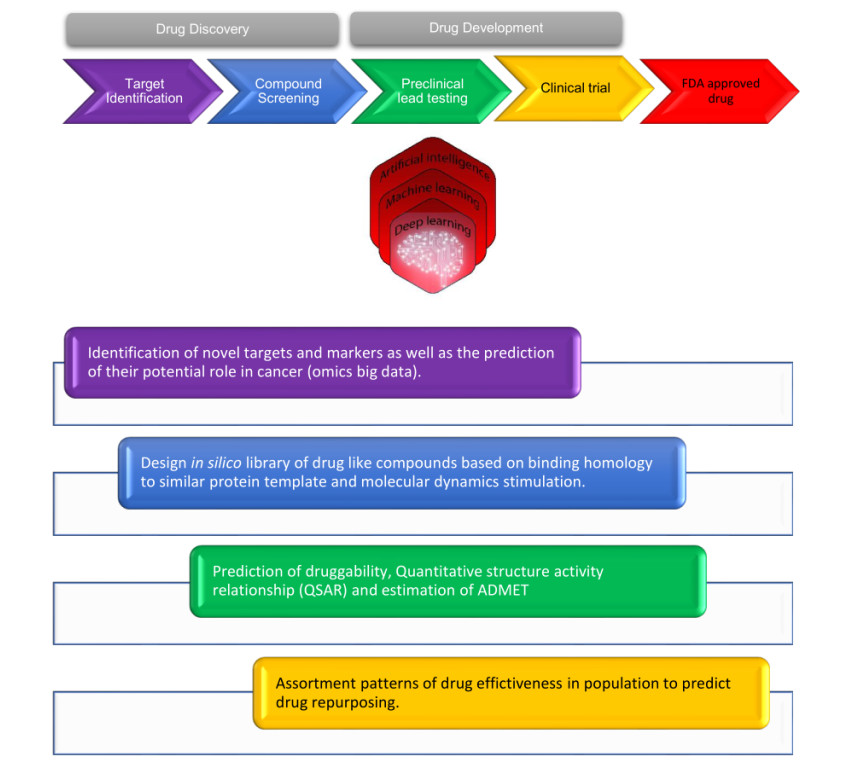
Figures(1) / Tables(1)

Rachael C. Adams, Behnam Rashidieh. Can computers conceive the complexity of cancer to cure it? Using artificial intelligence technology in cancer modelling and drug discovery[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6515-6530. doi: 10.3934/mbe.2020340







 DownLoad:
DownLoad: