Citation: Carl Graham, Jérôme Harmand, Sylvie Méléard, Josué Tchouanti. Bacterial metabolic heterogeneity: from stochastic to deterministic models[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5120-5133. doi: 10.3934/mbe.2020276

| [1] | J. Monod, Recherches Sur La Croissance Des Cultures Bactériennes, Hermann & Cie, Paris, 1942. |
| [2] | A. M. Liquori, A. Monroy, E. Parisi, A. Tripiciano, A theoretical equation for diauxic growth and its application to the kinetics of the early development of the sea urchin embryo, Differentiation, 20 (1981), 174-175. |
| [3] | V. Turon, Coupling Dark Fermentation with Microalgal Heterotrophy: Influence of Fermentation Metabolites Mixtures, Light, Temperature and Fermentation Bacteria on Microalgae Growth, Ph.D thesis, Université de Montpellier, 2015. |
| [4] | G. Van Dedem, M. Moo-Young, A model of diauxic growth, Biotechnol. Bioeng., 17 (1975), 1301-1312. |
| [5] | R. L. Bertrand, Lag phase is a dynamic, organized, adaptative, and evolvable period that prepares bacteria for cell division, J. Bacteriol., 201 (2019), e00697-18. |
| [6] | V. Takhaveev, M. Heinemann, Metabolic heterogeneity in clonal microbial populations, Curr. Opin. Microbiol., 45 (2018), 30-38. |
| [7] | B. Enjalbert, P. Millard, M. Dinciaux, J. C. Portais, F. Létisse, Acetate fluxes in Escherichia coli are determined by the thermodynamic contol of the Pta-AckA pathway, Sci. Rep., 7 (2017), 42135. |
| [8] | M. Barthe, J. Tchouanti, P. H. Gomes, C. Bideaux, D. Lestrade, C. Graham, et al., Duration of the glucose-xylose lag is controlled stochastically by the molecular switch XylR in Escherichia coli., Submitted, 2020. |
| [9] | S. N. Ethier, T. G. Kurtz, Markov Processes: Characterization and Convergence, John Wiley & Sons, 1986. |
| [10] | D. F. Anderson, T. G. Kurtz, Stochastic Analysis of Biochemical Systems, Springer, 2015. |
| [11] | V. Bansaye, S. Méléard, Stochastic Models for Structured Populations: Scaling Limits and Long Time Behavior, Springer, 2015. |
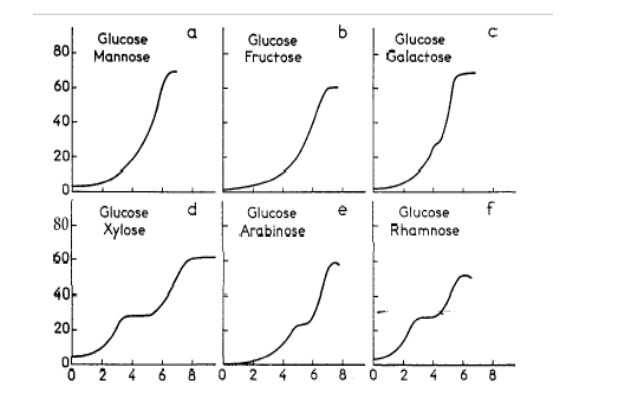
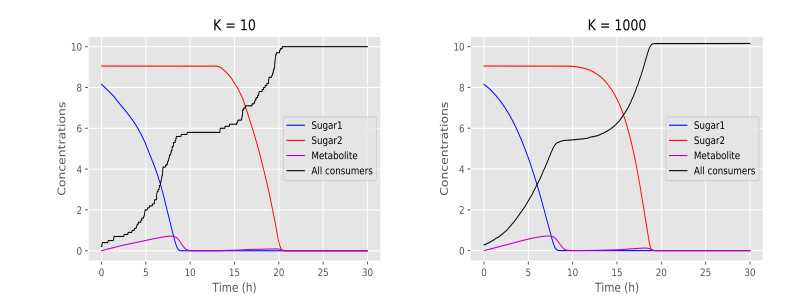
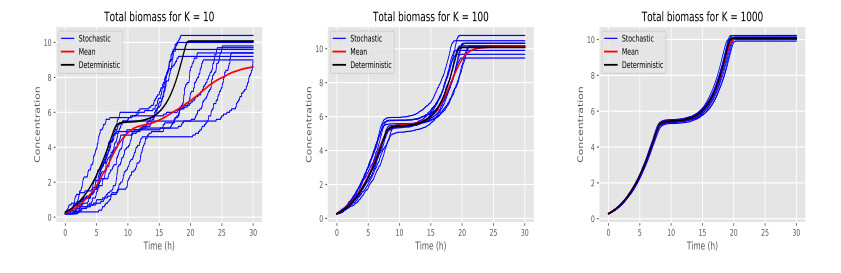
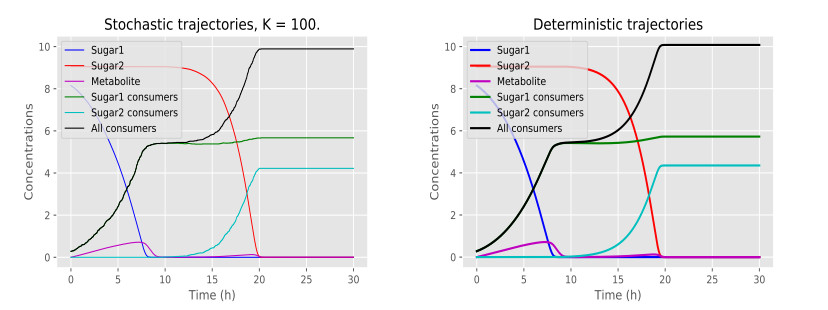
Figures(6) / Tables(1)

Carl Graham, Jérôme Harmand, Sylvie Méléard, Josué Tchouanti. Bacterial metabolic heterogeneity: from stochastic to deterministic models[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5120-5133. doi: 10.3934/mbe.2020276







 DownLoad:
DownLoad: