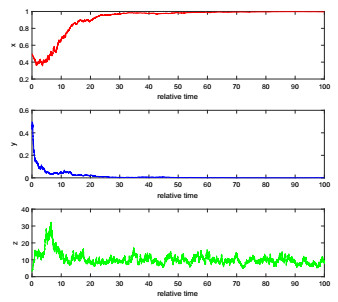
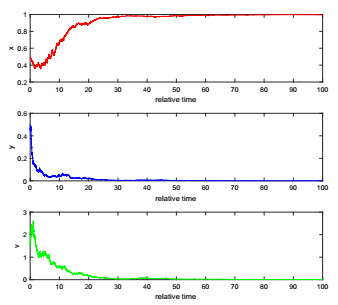
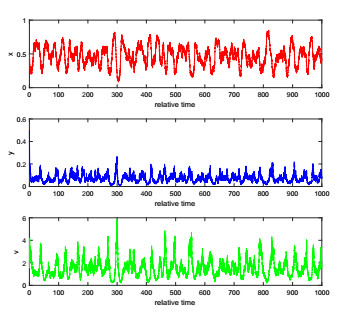
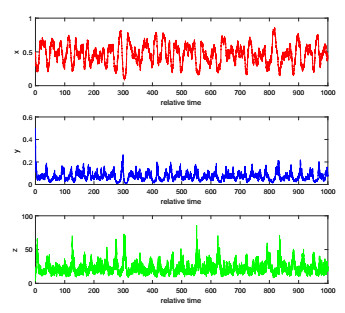
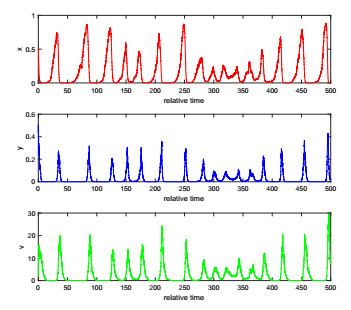
The complexity of oncolytic virotherapy arises from many factors. In this study, we incorporate environmental noise and stochastic effects to our basic deterministic model and propose a stochastic model for viral therapy in terms of Ito stochastic differential equations. We conduct a detailed analysis of the model using boundary methods. We find two combined parameters, one describes possibilities of eradicating tumors and one is an increasing function of the viral burst size, which serve as thresholds to classify asymptotical dynamics of the model solution paths. We show there are three ergodic invariant probability measures which correspond to equilibrium states of the deterministic model, and extra possibility to eradicate tumor due to strong variance of tumor growth rate and medium viral burst size. Numerical analysis demonstrates several typical solution paths with biological explanations. In addition, we provide some medical interpretations and implications.
Citation: Tuan Anh Phan, Jianjun Paul Tian. Basic stochastic model for tumor virotherapy[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4271-4294. doi: 10.3934/mbe.2020236
The complexity of oncolytic virotherapy arises from many factors. In this study, we incorporate environmental noise and stochastic effects to our basic deterministic model and propose a stochastic model for viral therapy in terms of Ito stochastic differential equations. We conduct a detailed analysis of the model using boundary methods. We find two combined parameters, one describes possibilities of eradicating tumors and one is an increasing function of the viral burst size, which serve as thresholds to classify asymptotical dynamics of the model solution paths. We show there are three ergodic invariant probability measures which correspond to equilibrium states of the deterministic model, and extra possibility to eradicate tumor due to strong variance of tumor growth rate and medium viral burst size. Numerical analysis demonstrates several typical solution paths with biological explanations. In addition, we provide some medical interpretations and implications.

| [1] | D. Cross, J. Burmester, Gene therapy for cancer treatment: Past, present, and future, Clin. Med. Res., 4 (2006), 218-227. |
| [2] |
X. M. Anguela, K. A. High, Entering the modern era of gene therapy, Annu. Rev. Med., 70 (2019), 273-288. doi: 10.1146/annurev-med-012017-043332

|
| [3] | E. A. Chiocca, Oncolytic viruses, Nat. Rev. Cancer, 2 (2002), 938-950. |
| [4] |
E. Kelly, S. J. Russell, History of oncolytic viruses: Genesis to genetic engineering, Mol. Ther., 15 (2007), 651-659. doi: 10.1038/sj.mt.6300108
|
| [5] | R. H. I. Andtbacka, H. L. Kaufman, F. Collichio, T. Amatruda, N. Senzer, J. Chesney, et al., Talimogene laherparepvec improves durable response rate in patients with advanced melanoma, J. Clin. Oncol., 33 (2015), 2780-2788. |
| [6] |
T. Liu, D. Kirn, Gene therapy progress and prospects cancer: oncolytic viruses, Gene Ther., 15 (2008), 877-884. doi: 10.1038/gt.2008.72
|
| [7] | D. Wodarz, Viruses as antitumor weapons: Defining conditions for tumor remission, Cancer Res., 61 (2001), 3501-3507. |
| [8] | D. Wodarz, Gene therapy for killing p53-negative cancer cells: Use of replicating versus nonreplicating agents, Hum. Gene Ther., 159 (2003), 153-159. |
| [9] |
D. Dingli, M. D. Cascino, K. Josic, S. J. Russell, Z. Bajzer, Mathematical modeling of cancer radiovirotherapy, Math. Biosci., 199 (2006), 55-78. doi: 10.1016/j.mbs.2005.11.001
|
| [10] |
Z. Bajzer, T. Carr, K. Josic, S. J. Russell, D. Dingli, Modeling of cancer virotherapy with recombinant measles viruses, J. Theor. Biol., 252 (2008), 109-22. doi: 10.1016/j.jtbi.2008.01.016
|
| [11] |
N. L. Komarova, D. Wodarz, ODE models for oncolytic virus dynamics, J. Theor. Biol., 263 (2010), 530-543. doi: 10.1016/j.jtbi.2010.01.009
|
| [12] |
J. P. Tian, The replicability of oncolytic virus: Defining conditions on tumor virotherapy, Math. Biosci. Eng., 8 (2011), 841-860. doi: 10.3934/mbe.2011.8.841
|
| [13] | T. A. Phan, J. P. Tian, The Role of the Innate Immune System in Oncolytic Virotherapy, Comput. Math. Methods Med., Volume 2017, Article ID 6587258, 17 pages. |
| [14] |
Y. Yuan, L. J. Allen, Stochastic models for virus and immune system dynamics, Math. Biosci., 234 (2011), 84-94. doi: 10.1016/j.mbs.2011.08.007
|
| [15] |
K. S. Kim, S. Kim, I. H. Jung, Dynamics of tumor virotherapy: A deterministic and stochastic model approach, Stoch. Anal. Appl., 34 (2016), 483-495. doi: 10.1080/07362994.2016.1150187
|
| [16] |
M. Rajalakshmi, M. Ghosh, Modeling treatment of cancer using virotherapy with generalized logistic growth of tumor cells, Stoch. Anal. Appl., 36 (2018), 1068-1086. doi: 10.1080/07362994.2018.1535319
|
| [17] |
M. Rajalakshmi, M. Ghosh, Modeling treatment of cancer using oncolytic virotherapy with saturated incidence, Stoch. Anal. Appl., 38 (2020), 565-579. doi: 10.1080/07362994.2019.1703743
|
| [18] | E. Allen, Modeling with Ito Stochastic Differential Equations, Springer, Dordrecht, The Netherlands, 2007. |
| [19] | J. Cresson, B. Puig, S. Sonner, Validating stochastic models: Invariance criteria for systems of stochastic differential equations and the selection of a stochastic Hodgkin-Huxley type model, Int. J. Biomath. Biostat., 2 (2013), 111-122. |
| [20] |
J. Cresson, B. Puig, S. Sonner, Stochastic models in biology and the invariance problem, Discrete Continuous Dyn. Syst. Ser. B, 21 (2016), 2145-2168. doi: 10.3934/dcdsb.2016041
|
| [21] |
J. Cresson, S. Sonner, A note on a derivation method for SDE models: Applications in biology and viability criteria, Stoch. Anal. Appl., 36 (2018), 224-239. doi: 10.1080/07362994.2017.1386571
|
| [22] | T. A. Phan, J. P. Tian, B. Wang, Dynamics of cholera epidemic models in fluctuating environments, Stoch. Dyn., (2020), In press. |
| [23] | A. Hening, H. D. Nguyen, Coexistence and extinction for stochastic Kolmogorov systems, Ann. Appl. Probab., 28 (2018), 1893-1942. |
| [24] | X. Mao, Stochastic differential equations and applications, 2nd edition, Woodhead Publishing Limited, 2007. |
| [25] | N. Ikeda, S. Watanabe, Stochastic Differential Equations and Diffusion Processes, 2nd edition, North-Holland Publishing Co., Amsterdam, 1989. |
| [26] | B. Jorgensen, Statistical Property of the Generalized Inverse Gaussian Distribution, SpringerVerlag New York, 1982. |
| [27] |
A. Friedman, J. P. Tian, G. Fulci, E. A. Chiocca, J. Wang, Glioma virotherapy: The effects of innate immune suppression and increased viral replication capacity, Cancer Res., 66 (2006), 2314-2319. doi: 10.1158/0008-5472.CAN-05-2661
|
Figures(7)

Tuan Anh Phan, Jianjun Paul Tian. Basic stochastic model for tumor virotherapy[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4271-4294. doi: 10.3934/mbe.2020236







 DownLoad:
DownLoad: