Citation: Zizi Wang, Zhiming Guo, Hal Smith. A mathematical model of oncolytic virotherapy with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1836-1860. doi: 10.3934/mbe.2019089

| [1] | R. Dunia and T. F. Edgar, Modeling of tumor growth undergoing virotherapy, Comput. Biol. Med., 41 (2011), 922–935. |
| [2] | S. J. Ries and C. H. Brandts, Oncolytic viruses for the treatment of cancer: current strategies and clinical trials, Drug Discov. Today, 9 (2004), 759–768. |
| [3] | M. J. Vähä-Koskela, J. E. Heikkilä and A. E. Hinkkanen, Oncolytic viruses in cancer therapy, Cancer Lett., 254 (2007), 178–216. |
| [4] | L. M. Wein, J. T. Wu, and D. H. Kirn, Validation and analysis of a mathematical model of a replication-competent oncolytic virus for cancer treatment, Cancer Res., 63 (2003), 1317–1324. |
| [5] | N. L. Komarova and D. Wodarz, ODE models for oncolytic virus dynamics, J. Theor. Biol., 263 (2010), 530–543. |
| [6] | K. Garber, China approves world's first oncolytic virus therapy for cancer treatment, J. Natl. Cancer Inst., 98 (2006), 298–300. |
| [7] | J. M. Markert, M. D. Medlock, S. D. Rabkin, et al., Conditionally replicating herpes simplex virus mutant, G207 for the treatment of malignant glioma: results of a phase I trial, Gene Ther., 7 (2000), 867–874. |
| [8] | L. K. Csatary, G. Gosztonyi, J. Szeberenyi, et al., MTH-68/H Oncolytic Viral Treatment in Human High-Grade Gliomas, J. Neuro-Oncol., 67 (2004), 83–93. |
| [9] | Y. Lin, H. Zhang, J. Liang, et al., Identification and characterization of alphavirus M1 as a selective oncolytic virus targeting ZAP-defective human cancers, Proc. Natl. Acad. Sci. USA, 111 (2014), 4504–4512. |
| [10] | D. Wodarz, Viruses as antitumor weapons, Cancer Res., 61 (2001), 3501–3507. |
| [11] | J. Heaney, T. Barrett, and S. L. Cosby, Inhibition of in vitro leukocyte proliferation by morbilliviruses, J. Virol., 76 (2002), 3579–3584. |
| [12] | Ž. Bajzer, T. Carr, K. Josi´c, et al., Modeling of cancer virotherapy with recombinant measles viruses, J. Theor. Biol., 252 (2008), 109–122. |
| [13] | D. Dingli, M. D. Cascino, K. Josi´c, et al., Mathematical modeling of cancer radiovirotherapy, Math. Biosci., 199 (2006), 55–78. |
| [14] | Y. Wang, J. P. Tian and J. Wei, Lytic cycle: A defining process in oncolytic virotherapy, Appl. Math. Model., 37 (2013), 5962–5978. |
| [15] | E. Beretta and Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144–1165. |
| [16] | B. R. Dix, S. J. O'Carroll, J. Colleen, et al., Efficient induction of cell death by adenoviruses requires binding of E1B55k and p53, Cancer Res., 60 (2000), 2666–2672. |
| [17] | A. R. Hall, B. R. Dix, S. J. O'Carroll, et al., p53-dependent cell death/apoptosis is required for a productive adenovirus infection, Nat. Med., 4 (1998), 1068–1072. |
| [18] | J. N. Harada and A. J. Berk, p53-Independent and -Dependent Requirements for E1B-55K in Adenovirus Type 5 Replication, J. Virol., 4 (1999), 5333–5344. |
| [19] | M. Ramachandra, A. Rahman, A. Zou, et al., Re-engineering adenovirus regulatory pathways to enhance oncolytic specificity and efficacy, Nat. Biotechnol., 19 (2001), 1035–1041. |
| [20] | G. Gonzalez-Parra, H. M. Dobrovolny, D. F. Aranda, et al., Quantifying rotavirus kinetics in the REH tumor cell line using in vitro data, Virus Res., 244 (2018), 53–63. |
| [21] | J. A. Borghans, R. J. de Boer and L. A. Segel, Extending the quasi-steady state approximation by changing variables, Bull. Math. Biol., 58 (1996), 43–63. |
| [22] | A. Friedman, J. P. Tian, G. Fulci, et al., Glioma virotherapy: Effects of innate immune suppression and increased viral replication capacity, Cancer Res., 66 (2006), 2314–2319. |
| [23] | K. Li, J. Liang, Y. Lin, et al., A classical PKA inhibitor increases the oncolytic effect of M1 virus via activation of exchange protein directly activated by cAMP 1, Oncotarget, 7 (2016), 48443– 48455. |
| [24] | V. L. de Rioja, N. Isern and J. Fort, A mathematical approach to virus therapy of glioblastomas, Biol. Direct, 11 (2016), 1. |
| [25] | J. T. Wu, D. H. Kirn and L. M. Wein, Analysis of a three-way race between tumor growth, a replication-competent virus and an immune response, Bull. Math. Biol., 66 (2004), 605–625. |
| [26] | H. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000), 599–653. |
| [27] | H. L. Smith, An introduction to delay differential equations with applications to the life sciences, Texts in Applied Mathematics. |
| [28] | H. L. Smith and H. R. Thieme, Dynamical systems and population persistence, American Mathematical Soc., 118 (2011). |
| [29] | J. T.Wu, H. M. Byrne, D. H. Kirn, et al., Modeling and analysis of a virus that replicates selectively in tumor cells, Bull. Math. Biol., 63 (2001), 731–768. |
| [30] | Z. Wang, Z. Guo and H. Peng, A mathematical model verifying potent oncolytic efficacy of M1 virus, Math. Biosci., 63 (2001), 731–768. |
| [31] | Z. Wang, Z. Guo and H. Peng, Dynamical behavior of a new oncolytic virotherapy model based on gene variation, Discrete Cont. Dyn. S., 10 (2017), 1079–1093. |
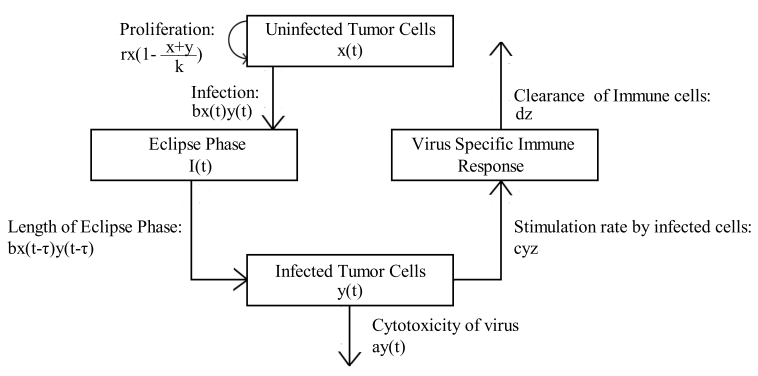
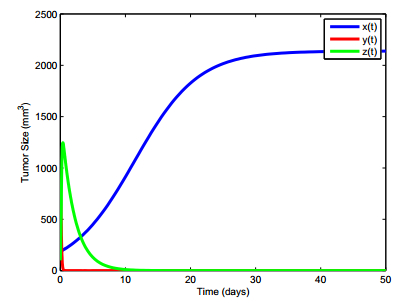
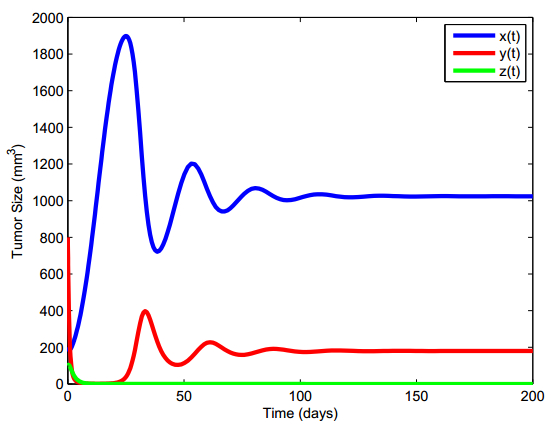
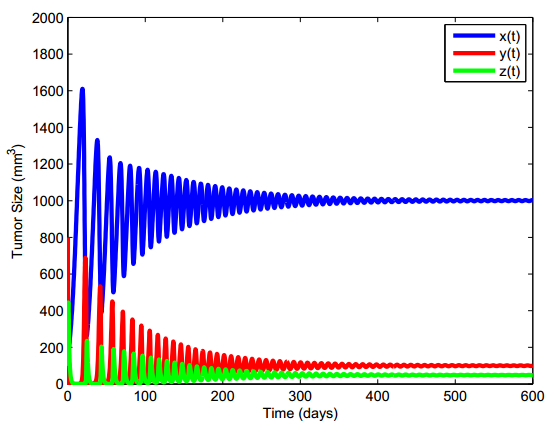
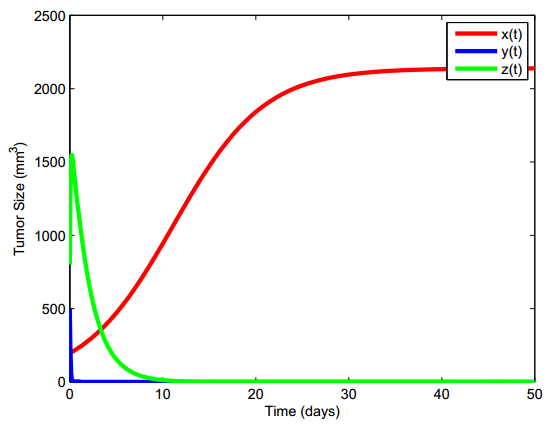
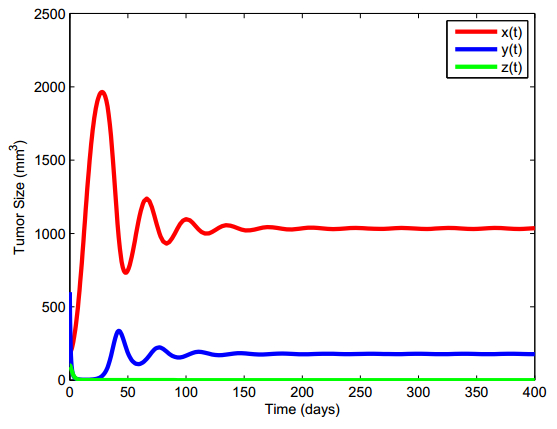
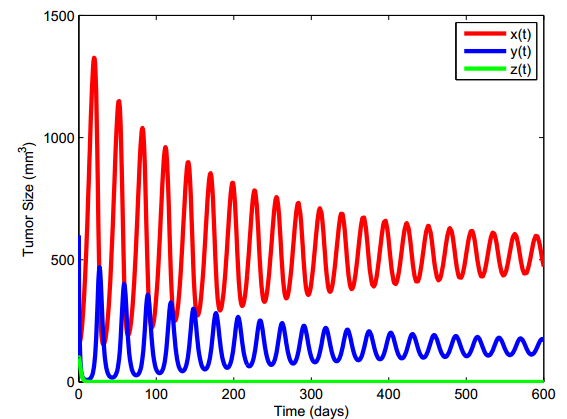
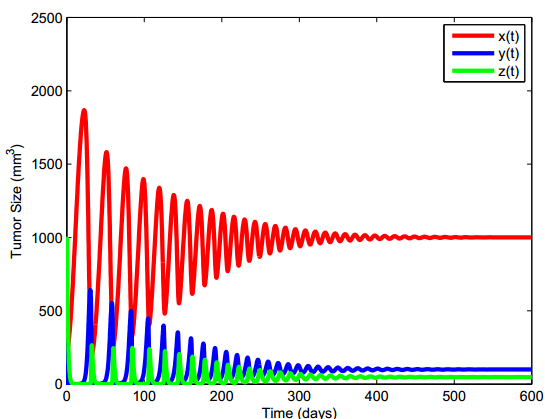
Figures(13) / Tables(5)

Zizi Wang, Zhiming Guo, Hal Smith. A mathematical model of oncolytic virotherapy with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1836-1860. doi: 10.3934/mbe.2019089







 DownLoad:
DownLoad: