Citation: Yousef Rohanizadegan, Stefanie Sonner, Hermann J. Eberl. Discrete attachment to a cellulolytic biofilm modeled by an Itô stochastic differential equation[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2236-2271. doi: 10.3934/mbe.2020119

| [1] | Z. Lewandowski, H. Beyenal, Fundamentals of Biofilm research, CRC Press, Boca Raton, 2007. |
| [2] | B. D'Acunto, L. Frunzo, V. Luongo, M. R. Mattei, Free boundary approach for the attachment in the initial phase of multispecies biofilm growth, Zeitschrift für angewandte Mathematik und Physik, 70 (2019), 91. |
| [3] | B. D'Acunto, L. Frunzo, M. R. Mattei, Continuum approach to mathematical modellingof multispecies biofilms, Ric. di Mat., 66 (2017), 153-169. |
| [4] | A. Masic, H. J. Eberl, Persistence in a single species cstr model with suspended flocs and wall attached biofilms, Bull. Math. Biol., 74 (2012), 1001-1026. |
| [5] | A. Masic, H. J. Eberl, A modeling and simulation study of the role of suspended microbial populations in nitrification in a biofilm reactor, Bull. Math. Biol., 76 (2014), 27-58. |
| [6] | H. J. Gaebler, H. J. Eberl, A simple model of biofilm growth in a porous medium that accounts for detachment and attachment of suspended biomass and their contribution to substrate degradation, Eur. J. Appl. Math., 29 (2018), 1110-1140. |
| [7] | E. Alkvist, I. Klapper, A multidimensional multispecies continuum modelfor heterogeneous biofilm development, Bull. Math. Biol., 69 (2007), 765-789. |
| [8] | B. V. Merkey, B. E. Rittmann, D. L. Chopp, Modeling how soluble microbial products (smp) support heterotrophic bacteria in autotroph-based biofilms, J. Theor. Biol., 259 (2009), 670-683. |
| [9] | N. G. Cogan, Two-fluid model of biofilm disinfection, Bull. Math. Biol., 70 (2008), 800-819. |
| [10] | J. Dockery, I. Klapper, Finger formation in biofilms, SIAM J. Appl. Math., 62 (2002), 853-869. |
| [11] | J. P. Ward, J. R. King, Thin-film modelling of biofilm growth and quorum sensing, J. Eng. Math., 73 (2012), 71-92. |
| [12] | H. J. Eberl, D. F. Parker, M. Van Loosdrecht, A new deterministic spatio-temporal continuum model for biofilm development, Comput. Math. Methods Med., 3 (2001), 161-175. |
| [13] | K. A. Rahman, R. Sudarsan, H. J. Eberl, A mixed-culture biofilm model with cross-diffusion, Bull. Math. Biol., 77 (2015), 2086-2124. |
| [14] | M. Whidden, N. Cogan, M. Donahue, F. Navarrete, L. D. L. Fuente, A two-dimensional multiphase model of biofilmformation in microfluidic chambers, Bull. Math. Biol., 77 (2015), 2161-2179. |
| [15] | F. El-Moustaid, A. Eladdadi, L. Uys, Modeling bacterial attachment to surfaces at and early stage of biofilm development, Math. Biosci. Eng., 10 (2013), 821-841. |
| [16] | H. J. Eberl, H. Khassehkhan, L. Demaret, A mixed-culture model of a probiotic biofilm control system, Comput. Math. Methods Med., 11 (2010), 99-118. |
| [17] | A. Carroll, C. Somerville, Cellulosic biofuels, Annu. Rev. Plant Biol., 60 (2009), 165-182. |
| [18] | M. H. Langholtz, B. J. Stokes, L. M. Eaton, 2016 billion-ton report: Advancing domestic resources for a thriving bioeconomy, available from: https://bit.ly/2LLjdBS, 2016, URL https://doi.org/10.2172/1271651, (accessed 12-June-2018). |
| [19] | J. G. Linger, A. Darzins, Consolidated Bioprocessing, 267-280, Springer New York, New York, NY, 2013. |
| [20] | B. G. Schuster, M. S. Chinn, Consolidated bioprocessing of lignocellulosic feedstocks for ethanol fuel production, BioEnergy Res., 6 (2013), 416-435. |
| [21] | A. Dumitrache, G. Wolfaardt, G. Allen, S. N. Liss, L. R. Lynd, Form and function of Clostridium thermocellum biofilms, Appl. Environ. Microbiol., 79 (2013), 231-239. |
| [22] | V. Mbaneme-Smith, M. S. Chinn, Consolidated bioprocessing for biofuel production: recent advances, Energy Emission Control Technol., 3 (2015), 23-44. |
| [23] | Z.-W. Wang, S.-H. Lee, J. G. Elkins, J. L. Morrell-Falvey, Spatial and temporal dynamics of cellulose degradation and biofilm formation by caldicellulosiruptor obsidiansis and clostridium thermocellum, AMB Express, 1 (2011), 30. |
| [24] | M. van Loosdrecht, J. Heijnen, H. Eberl, J. Kreft, C. Picioreanu, Mathematical modelling of biofilm structures, Antonie van Leeuwenhoek, 81 (2002), 245-256. |
| [25] | D. G. Davies, M. R. Parsek, J. P. Pearson, B. H. Iglewski, J. W. Costerton, E. P. Greenberg, The involvement of cell-to-cell signals in the development of a bacterial biofilm, Science, 280 (1998), 295-298. |
| [26] | S. M. Hunt, E. M. Werner, B. Huang, M. A. Hamilton, P. S. Stewart, Hypothesis for the role of nutrient starvation in biofilm detachment, Appl. Environ. Microbiol., 70 (2004), 7418-7425. |
| [27] | J. Zhu, J. J. Mekalanos, Quorum sensing-dependent biofilms enhance colonization in vibrio cholerae, Dev. Cell, 5 (2003), 647-656. |
| [28] | Y. H. An, R. J. Friedman, Concise review of mechanisms of bacterial adhesion to biomaterial surfaces, J. Biomed. Mater. Res., 43 (1998), 338-348. |
| [29] | H. Eberl, E. Jalbert, A. Dumitrache, G. Wolfaardt, A spatially explicit model of inverse colony formation of cellulolytic biofilms, Biochem. Eng. J., 122 (2017), 141-151. |
| [30] | P. E. Kloeden, E. Platen, Numerical Solution of Stochastic Differential Equations, Springer-Verlag, Berlin Heidelberg, 1992. |
| [31] | B. Øksendal, Stochastic differential equations: An introduction with applications, Springer, Berlin Heidelberg, 2003. |
| [32] | A. Dumitrache, G. Wolfaardt, G. Allen, S. N. Liss, L. R. Lynd, Tracking the cellulolytic activity of Clostridium thermocellum biofilms, Biotech. Biofuels, 6 (2013), 175. |
| [33] | G. J. Lord, C. E. Powell, T. Shardlow, An introduction to computational stochastic PDEs, 50, Cambridge University Press, New York, NY, 2014. |
| [34] | J. Cresson, M. Efendiev, S. Sonner, On the positivity of solutions of systems of stochastic pdes, ZAMM - J. Appl. Math. Mech. / Zeitschrift für Angewandte Mathematik und Mechanik, 93 (2013), 414-422. |
| [35] | J. Cresson, B. Puig, S. Sonner, Stochastic models in biology and the invariance problem, Discrete Cont. Dyn- B, 21 (2016), 2145-2168. |
| [36] | R. Fox, Stochastic versions of the Hodgkin-Huxley equations, Biophys. J., 72 (1997), 2068-2074. |
| [37] | A. Kamina, R. W. Makuch, H. Zhao, A stochastic modeling of early HIV-1 population dynamics, Math. Biosci., 170 (2001), 187-198. |
| [38] | H. C. Tuckwell, E. L. Corfec, A stochastic model for early hiv-1 population dynamics, J. Theor. Biol., 195 (1998), 451-463. |
| [39] | H. J. Eberl, L. Demaret, A finite difference scheme for a degenerated diffusion equation arising in microbial ecology, Electron. J. Differ. Equ, 15 (2007), 77-95. |
| [40] | A. Dumitrache, H. J. Eberl, D. G. Allen, G. M. Wolfaardt, Mathematical modeling to validate on-line co2 measurements as a metric for cellulolytic biofilm activity in continuous-flow bioreactors, Biochem. Eng. J., 101 (2015), 55-67. |
| [41] | M. A. Efendiev, S. V. Zelik, H. J. Eberl, Existence and longtime behavior of a biofilm model, Commun. Pure Appl. Anal., 8 (2009), 509-531. |
| [42] | M. Ghasemi, H. J. Eberl, Time adaptive numerical solution of a highly degenerate diffusion-reaction biofilm model based on regularisation, J. Sci. Comput., 74 (2018), 1060-1090. |
| [43] | J. Cresson, B. Puig, S. Sonner, Validating stochastic models: invariance criteria for systems of stochastic differential equations and the selection of a stochastic Hodgkin-Huxley type model, Internat. J. Biomath. Biostat., 2 (2013), 111-222. |
| [44] | X. Han, P. E. Kloeden, Random Ordinary Differential Equations and Their Numerical Solution, Springer, Singapore, 2017. |
| [45] | P. E. Kloeden, E. Platen, H. Schurz, Numerical Solution of SDE Through Computer Experiments, Springer-Verlag, Berlin Heidelberg, 1994. |
| [46] | J. Monod, The growth of bacterial cultures, Ann. Rev. Microbiol., 3 (1949), 371-394. |
| [47] | Y. Rohanizadegan, A Stochastic Formulation of Bacterial Attachment in a Spatially Explicit Model of Cellulolytic Biofilm Formation, Master's thesis, University of Guelph, Canada, 2018, available from: URL https://atrium.lib.uoguelph.ca/xmlui/handle/10214/14673. |
| [48] | W. Walter, Ordinary Differential Equations, Springer New York, 1998. |
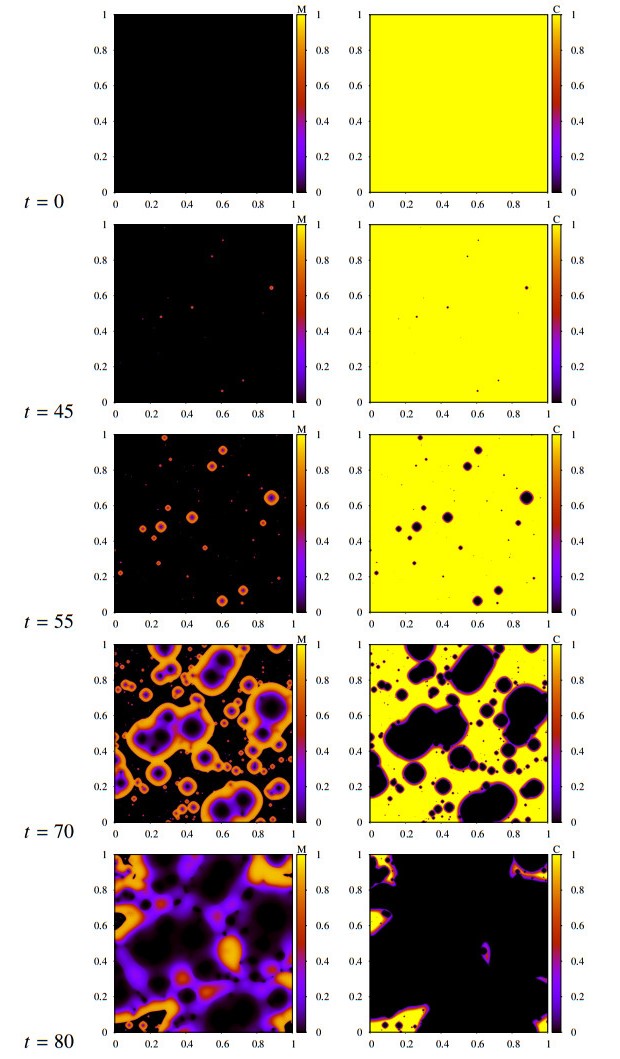
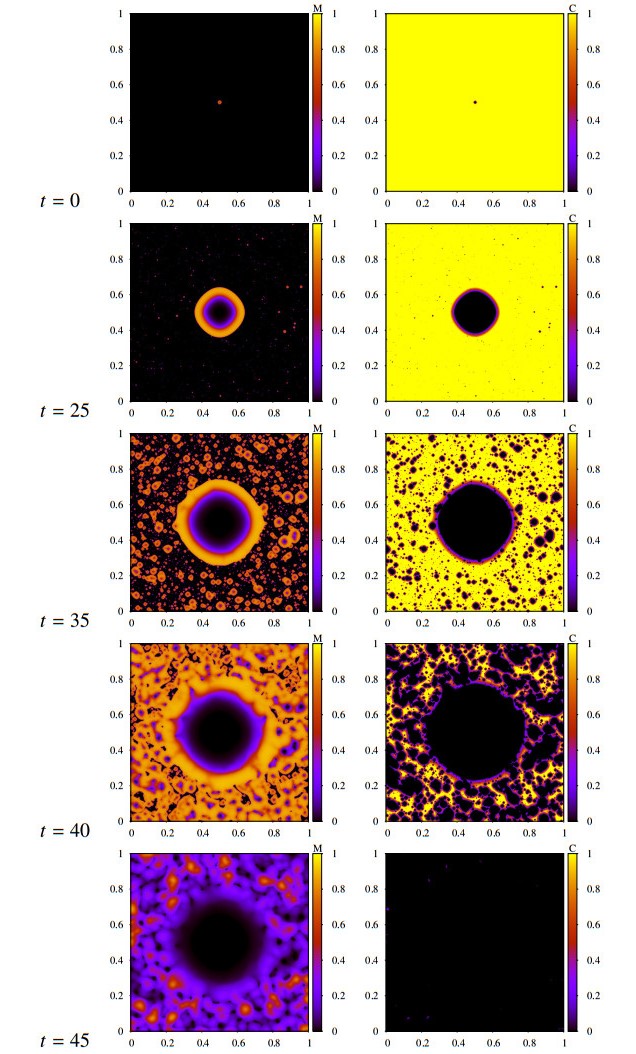
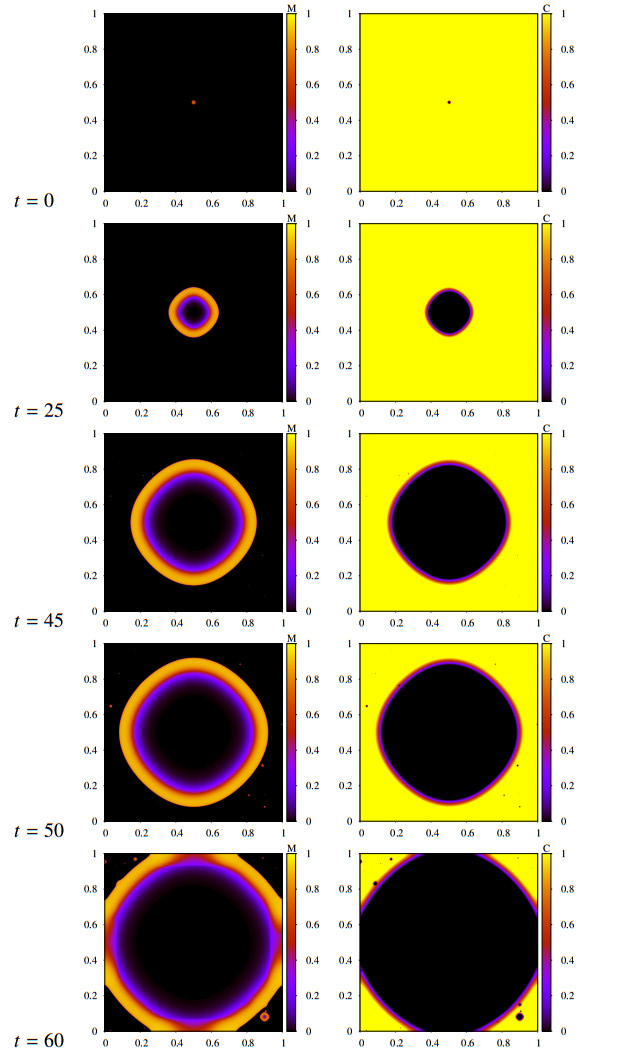
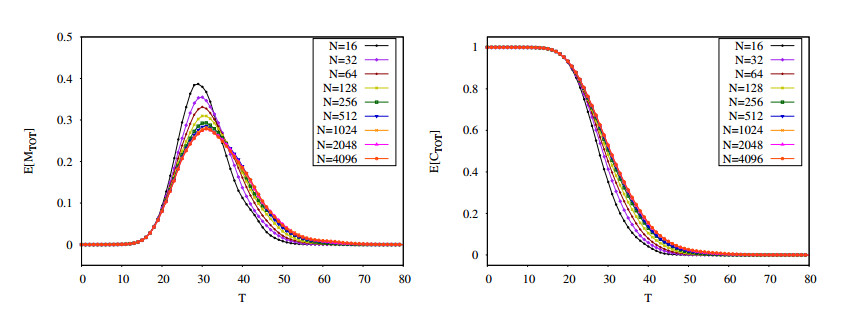
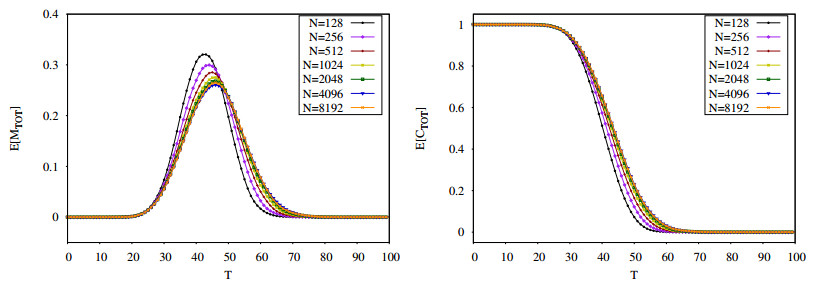
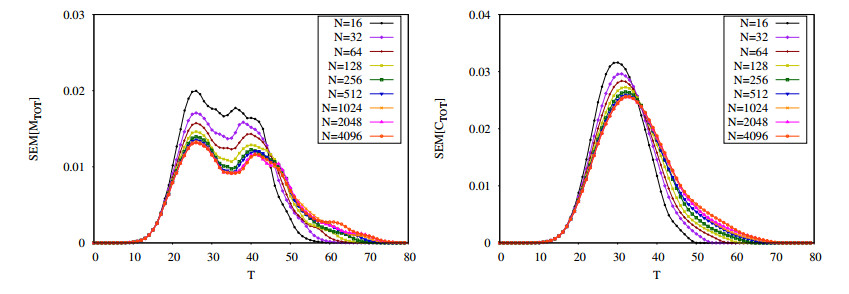
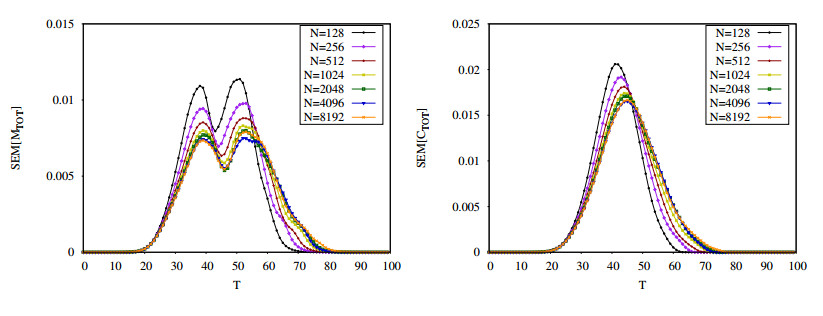
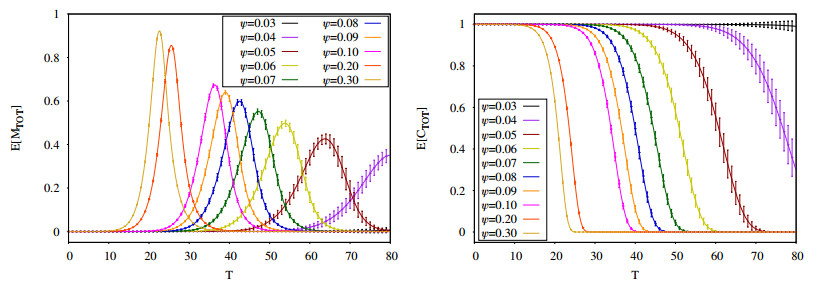
Figures(16) / Tables(2)

Yousef Rohanizadegan, Stefanie Sonner, Hermann J. Eberl. Discrete attachment to a cellulolytic biofilm modeled by an Itô stochastic differential equation[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2236-2271. doi: 10.3934/mbe.2020119







 DownLoad:
DownLoad: