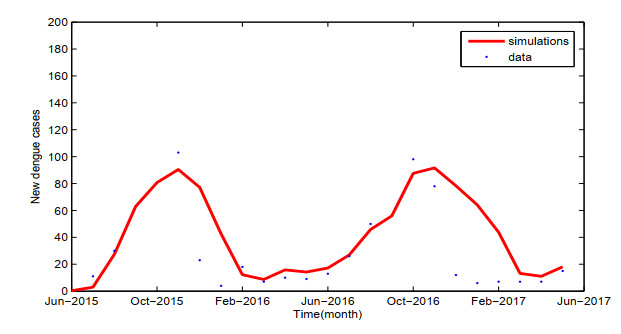
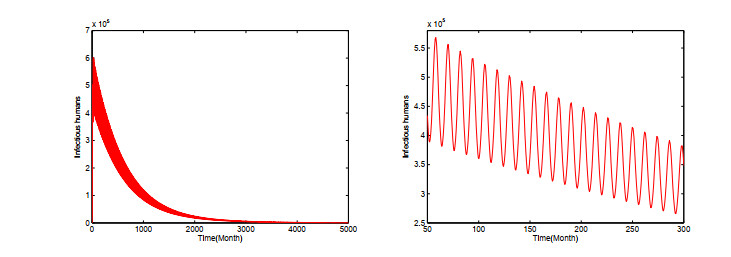
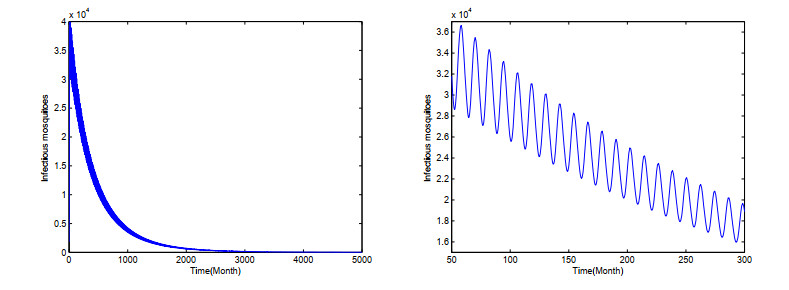
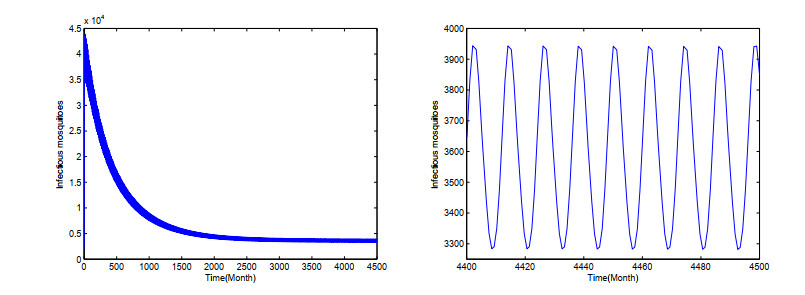
Dengue fever is a re-emergent mosquito-borne disease, which prevails in tropical and subtropical regions, mainly in urban and peri-urban areas. Its incidence has increased fourfold since 1970, and dengue fever has become the most prevalent mosquito-borne disease in humans now. In order to study the effect of temperature on the dengue virus transmission, we formulate a dengue virus transmission model with maturation delay for mosquito production and seasonality. The basic reproduction number $\mathbb{R}_0$ of the model is computed, and results suggest that the dengue fever will die out if $\mathbb{R}_0$ < 1, and there exists at least one positive periodic solution and the disease will persist if $\mathbb{R}_0$ > 1. Theoretical results are applied to the outbreak of dengue fever in Guangdong province, China. Simulations reveal that the temperature change causes the periodic oscillations of dengue fever cases, which is good accordance with the reported cases of dengue fever in Guangdong province. Our study contributes to a better understanding of dengue virus transmission dynamics and proves beneficial in preventing and controlling of dengue fever.
Citation: Haitao Song, Dan Tian, Chunhua Shan. Modeling the effect of temperature on dengue virus transmission with periodic delay differential equations[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4147-4164. doi: 10.3934/mbe.2020230
Dengue fever is a re-emergent mosquito-borne disease, which prevails in tropical and subtropical regions, mainly in urban and peri-urban areas. Its incidence has increased fourfold since 1970, and dengue fever has become the most prevalent mosquito-borne disease in humans now. In order to study the effect of temperature on the dengue virus transmission, we formulate a dengue virus transmission model with maturation delay for mosquito production and seasonality. The basic reproduction number $\mathbb{R}_0$ of the model is computed, and results suggest that the dengue fever will die out if $\mathbb{R}_0$ < 1, and there exists at least one positive periodic solution and the disease will persist if $\mathbb{R}_0$ > 1. Theoretical results are applied to the outbreak of dengue fever in Guangdong province, China. Simulations reveal that the temperature change causes the periodic oscillations of dengue fever cases, which is good accordance with the reported cases of dengue fever in Guangdong province. Our study contributes to a better understanding of dengue virus transmission dynamics and proves beneficial in preventing and controlling of dengue fever.

| [1] | T. Bancroft, On the etiology of dengue fever, Austral. Med. Gaz., 25 (1906), 17-18. |
| [2] | S. Halstead, Dengue, Lancet, 370 (2007), 1644-1652. |
| [3] |
I. Kautner, M. Robinson, U. Kuhnle, Dengue virus infection: Epidemiology, pathogenesis, clinical presentation, diagnosis, and prevention, J. Pediatr., 131 (1997), 516-524. doi: 10.1016/S0022-3476(97)70054-4

|
| [4] |
T. Senaratne, F. Noordeen, Diagnosis of dengue in Sri Lanka: Improvements to the existing state of the art in the island, Trans. R. Soc. Trop. Med. Hyg., 108 (2014), 685-691. doi: 10.1093/trstmh/tru131
|
| [5] |
C. Pagliari, J. Quaresma, E. Fermandes, F. Stegun, R. Brasil, H. de Andrade Jr, et al., Immunopathogenesis of dengue hemorrhagic fever: Contribution to the study of human liver lesions, J. Med. Virol., 86 (2014), 1193-1197. doi: 10.1002/jmv.23758
|
| [6] | Public Health Agency of Canada. Available from: https://www.canada.ca/en/public-health.html. |
| [7] | Dengue and severe dengue, World Health Organization, 2020. Available from: https://www.who.int/en/news-room/fact-sheets/detail/dengue-and-severe-dengue. |
| [8] |
J. Blaney, N. Sathe, C. Hanson, C. Firestone, B. Murphy, S. Whitehead, Vaccine candidates for dengue virus type 1 (DEN1) generated by replacement of the structural genes of rDEN4 and rDEN4Δ30 with those of DEN1, Virol. J., 4 (2007), 23. doi: 10.1186/1743-422X-4-23
|
| [9] |
S. Sang, B. Chen B, H. Wu, Z. Yang, B. Di, L. Wang, et al., Dengue is still an imported disease in China: A case study in Guangzhou, Infect. Genet. Evol., 32 (2015), 178-190. doi: 10.1016/j.meegid.2015.03.005
|
| [10] |
L. Esteva, C. Vargas, Analysis of a dengue disease transmission model, Math. Biosci., 150 (1998), 131-151. doi: 10.1016/S0025-5564(98)10003-2
|
| [11] |
L. Esteva, C. Vargas, Influence of vertical and mechanical transmission on the dynamics of dengue disease, Math. Biosci., 167 (2000), 51-64. doi: 10.1016/S0025-5564(00)00024-9
|
| [12] |
G. Chowell, P. Diaz-Duenas, J. Miller, A. Alcazar-Velazco, J. Fenimore, C. Castillo-Chavez, Estimation of the reproduction number of dengue fever from spatial epidemic data, Math. Biosci., 208 (2007), 571-589. doi: 10.1016/j.mbs.2006.11.011
|
| [13] | A. Khan, M. Hassan, M. Imran, Estimating the basic reproduction number for single-strain dengue fever epidemics, Infect. Dise. Poverty, 1 (2014), 12. |
| [14] |
J. Tewa, J. Dimi, S. Bowong, Lyapunov functions for a dengue disease transmission model, Chaos, Solitons Fractals, 39 (2009), 936-941. doi: 10.1016/j.chaos.2007.01.069
|
| [15] | H. Yang, C. Ferreira, Assessing the effects of vector control on dengue transmission, Appl. Math. Comput., 198 (2008), 401-413. |
| [16] |
S. Garba, A. Gumel, Backward bifurcations in dengue transmission dynamics, Math. Biosci., 215 (2008), 11-25. doi: 10.1016/j.mbs.2008.05.002
|
| [17] |
A. Abdelrazec, J. Belair, C. Shan, H. Zhu, Modeling the spread and control of dengue with limited public health resources, Math. Biosci., 271 (2016), 136-145. doi: 10.1016/j.mbs.2015.11.004
|
| [18] |
S. Chen, C. Liao, C. Chio, H. Chou, S. You, Y. Cheng, Lagged temperature effect with mosquito transmission potential explains dengue variability in southern Taiwan: insights from a statistical analysis, Sci. Total Environ., 408 (2010), 4069-4075. doi: 10.1016/j.scitotenv.2010.05.021
|
| [19] |
C. Shan, H. Zhu, Bifurcations and complex dynamics of an SIR model with the impact of the number of hospital beds, J. Differ. Equ., 257 (2014), 1662-1688. doi: 10.1016/j.jde.2014.05.030
|
| [20] |
L. Rueda, K. Patel, R. Axtell, R. Stinner, Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae), J. Med. Entomol., 27 (1990), 892-898. doi: 10.1093/jmedent/27.5.892
|
| [21] |
W. Tun-Lin, T. Burkot, B. Kay, Effects of temperature and larval diet on development rates and survival of the dengue vector Aedes aegypti in north Queensland, Australia, Med. Vet. Entomol., 14 (2000), 31-37. doi: 10.1046/j.1365-2915.2000.00207.x
|
| [22] |
M. Tong, A. Hansen, S. Hanson-Easey, J. Xiang, S. Cameron, Q. Liu, et al., Perceptions of capacity for infectious disease control and prevention to meet the challenges of dengue fever in the face of climate change: A survey among CDC staff in Guangdong Province, China, Environ. Res., 148 (2016), 295-302. doi: 10.1016/j.envres.2016.03.043
|
| [23] |
G. Fan, J. Liu, P. Van den Driessche, J. Wu, H. Zhu, The impact of maturation delay of mosquitoes on the transmission of West Nile virus, Math. Biosci., 228 (2010), 119-126. doi: 10.1016/j.mbs.2010.08.010
|
| [24] | M. Li, G. Sun, L. Yakob, H. Zhu, Z. Jin, W. Zhang, The driving force for 2014 dengue outbreak in Guangdong, China, PloS One, 11 (2016), e0166211. |
| [25] |
L. Bai, L. Morton, Q. Liu, Climate change and mosquito-borne diseases in China: A review, Glob. Health, 9 (2013), 10. doi: 10.1186/1744-8603-9-10
|
| [26] | J. Gubler, The Arboviruses: Epidemiology and Ecology, Ⅱ. CRC Press, Florida, 1989. |
| [27] |
C. Bowman, A. Gumel, P. Van den Driessche, J. Wu, H. Zhu, A mathematical model for assessing control strategies against West Nile virus, Bull. Math. Biol., 67 (2005), 1107-1133. doi: 10.1016/j.bulm.2005.01.002
|
| [28] |
Y. Lou, X. Zhao, A climate-based malaria transmission model with structured vector population, SIAM J. Appl. Math., 70 (2010), 2023-2044. doi: 10.1137/080744438
|
| [29] |
H. Wan, H. Zhu, A new model with delay for mosquito population dynamics, Math. Biosci. Eng., 11 (2014), 1395-1410. doi: 10.3934/mbe.2014.11.1395
|
| [30] |
X. Wang, Q. Zhao, A periodic vector-bias malaria model with incubation period, SIAM J. Appl. Math., 77 (2017), 181-201. doi: 10.1137/15M1046277
|
| [31] |
K. Cooke, P. Van den Driessche, X. Zou, Interaction of maturation delay and nonlinear birth in population and epidemic models, J. Math. Biol., 39 (1999), 332-352. doi: 10.1007/s002850050194
|
| [32] |
J. Velascohernandez, A model for Chagas disease involving transmission by vectors and blood transfusion, Theor. Popul. Biol., 46 (1994), 1-31. doi: 10.1006/tpbi.1994.1017
|
| [33] | Y. Lou, X. Zhao, Threshold dynamics in a time-delayed periodic SIS epidemic model, Discrete Cont. Dyn. B., 126 (2009), 169-186. |
| [34] |
W. Wang, Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Differ. Equ., 20 (2008), 699-717. doi: 10.1007/s10884-008-9111-8
|
| [35] |
D. Xu, Q. Zhao, Dynamics in a periodic competitive model with stage structure, J. Math. Anal. Appl., 311 (2005), 417-438. doi: 10.1016/j.jmaa.2005.02.062
|
| [36] | H. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Society, 2008. |
| [37] | T. Kato, Perturbation Theory for Linear Operators, Springer Science and Business Media, 2013. |
| [38] |
F. Zhang, Q. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007), 496-516. doi: 10.1016/j.jmaa.2006.01.085
|
| [39] | Q. Zhao, J. Borwein, P. Borwein, Dynamical Systems in Population Biology, Springer, New York, 2017. |
| [40] |
P. Magal, X. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM J. Math. Anal., 37 (2005), 251-275. doi: 10.1137/S0036141003439173
|
| [41] | E. Chikaki, H. Ishikawa, A dengue transmission model in Thailand considering sequential infections with all four serotypes, J. Infect. Dev. Ctries., 3 (2009), 711-722. |
| [42] |
M. Andraud, N. Hens, C. Marais, P. Beutels, Dynamic epidemiological models for dengue transmission: A systematic review of structural approaches, PloS One, 7 (2012), e49085. doi: 10.1371/journal.pone.0049085
|
| [43] |
Z. Feng, J. Velasco-Hernandez, Competitive exclusion in a vector-host model for the dengue fever, J. Math. Biol., 35 (1997), 523-544. doi: 10.1007/s002850050064
|
| [44] |
H. Yang, M. Ferreira, K. Galvani, M. Andrighetti, D. Wanderley, Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue, Epidemiol. Infect., 137 (2009), 1188-1202. doi: 10.1017/S0950268809002040
|
| [45] |
D. Watts, D. Burke, B. Harrison, R. Whitmire, A. Nisalak, Effect of temperature on the vector efficiency of Aedes aegypti for dengue 2 virus, Am. J. Trop. Med. Hyg., 36 (1987), 143-152. doi: 10.4269/ajtmh.1987.36.143
|
| [46] |
S. Hales, N. De Wet, J. Maindonald, A. Woodward, Potential effect of population and climate changes on global distribution of dengue fever: An empirical model, Lancet, 360 (2002), 830- 834. doi: 10.1016/S0140-6736(02)09964-6
|
| [47] |
C. Shan, G. Fan, H. Zhu, Periodic phenomena and driving mechanisms in transmission of West Nile virus with maturation time, J. Dyn. Differ. Equ., 32 (2020), 1003-1026. doi: 10.1007/s10884-019-09758-x
|
| [48] | X. Li, J. Cao, An impulsive delay inequality involving unbounded time-varying delay and applications, IEEE Trans. Automat.Contr., 7 (2017), 3618-3625. |
| [49] | X. Li, X. Yang, T. Huang, Persistence of delayed cooperative models: Impulsive control method, Appl. Math. Comput., 342 (2019), 130-146. |

Figures(6) / Tables(2)

Haitao Song, Dan Tian, Chunhua Shan. Modeling the effect of temperature on dengue virus transmission with periodic delay differential equations[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4147-4164. doi: 10.3934/mbe.2020230







 DownLoad:
DownLoad: