Citation: Cameron J. Browne, Chang-Yuan Cheng. Age-structured viral dynamics in a host with multiple compartments[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 538-574. doi: 10.3934/mbe.2020029

| [1] | R. V. Culshaw, S. Ruan and G. Webb, A mathematical model of cell-to-cell spread of HIV-1 that includes a time delay, J. Math. Biol., 46 (2003), 425-444. |
| [2] | M. A. Nowak and R. M. May, Virus Dynamics, Oxford University Press, New York, 2000. |
| [3] | A. S. Perelson, D. E. Kirschner and R. de Boer, Dynamics of HIV infection of CD4+ T-cells, Math. Biosci., 114 (1993), 81-125. |
| [4] | C. Y. Cheng, Y. Dong and Y. Takeuchi, An age-structured virus model with two routs of infection in heterogeneous environments, Nonlinear Anal. RWA, 39 (2018), 464-491. |
| [5] | S. Nakaoka, S. Iwami and K. Sato, Dynamics of HIV infection in lymphoid tissue network, J. Math. Biol., 72 (2016), 909-938. |
| [6] | S. Wang, J. Wu and L. Rong, A note on the global properties of an age-structured viral dynamic model with multiple target cell populations, Math. Biosci. Eng., 14 (2017), 805-820. |
| [7] | M. C. Strain, D. D. Richman, J. K. Wong, et al., Spatiotemporal dynamics of HIV propagation, J. Theor. Biol., 218 (2002), 85-96. |
| [8] | S. I. Fox, Human Physiology, 7ed, Mcgraw-Hill, 2002. |
| [9] | V. Bronte and M. J. Pittet, The spleen in local and systemic regulation of immunity, Immunity, 39 (2013), 806-818. |
| [10] | C. Casteleyn, P. Cornillie, C. Van Ginneken, et al., Lymph drainage from the ovine tonsils: an anatomical study of the tonsillar lymph vessels, Anat. Histol. Embryol., 43 (2014), 482-489. |
| [11] | M. A. Weinreich and K. A. Hogquist, Thymic emigration: when and how T cells leave home, J. Immunol., 181 (2008), 2265-2270. |
| [12] | E. L. Haseltine, V. Lam, J. Yin, et al., Image-guided modeling of virus growth and spread, Bull. Math. Biol., 70 (2008), 1730-1748. |
| [13] | T. T. Murooka, M. Deruaz, F. Marangoni, et al., HIV-infected T cells aremigratory vehicles for viral dissemination, Nature, 490 (2012), 283-287. |
| [14] | G. A. Funk, V. A. Jansen, S. Bonhoeffer, et al., Spatial models of virus-immune dynamics, J. Theor. Biol., 233 (2005), 221-236. |
| [15] | W. Hübner, G. P. McNerney, P. Chen, et al., Quantitative 3D video microscopy of HIV transfer across T cell virological synapses, Science, 323 (2009), 1743-1747. |
| [16] | C. Zhang, S. Zhou, E. Groppelli, et al., Hybrid spreading mechanisms and T cell activation shape the dynamics of HIV-1 infection, PLOS Computational Biology, 11 (4) (2015), e1004179. |
| [17] | H. R. Thieme and C. Castillo-Chavez, How may the infection-age-dependent infectivity affect the dynamics of HIV/AIDS?, SIAM J. Appl. Math., 53 (1993), 1447-1479. |
| [18] | P. W. Nelson, M. A. Gilchrist, D. Coombs, et al., An age-structured model of HIV infection that allows for variations in the production rate of viral particles and the death rate of productively infected cells, Math. Biosci. Eng., 9 (2004), 267-288. |
| [19] | P. Magal, C. C. McCluskey and G. F. Webb, Lyapunov functional and global asymptotic stability for an infection-age model, Appl. Anal., 89 (2010), 1109-1140. |
| [20] | T. Kuniya, J. Wang and H. Inaba, A multi-group SIR epidemic model with age structure, Discrete Contin. Dyn. Syst. Ser. B, 21 (2016), 3515-3550. |
| [21] | J. Yang, Y. Chen and F. Xu, Effect of infection age on an SIS epidemic model on complex networks, J. Math. Biol., 73 (2016), 1227-1249. |
| [22] | P. Magal and C. C. McCluskey, Two-group infection age model including an application to nosocomial infection, SIAM J. Appl. Math., 73 (2013), 1058-1095. |
| [23] | H. R. Thieme, Semiflows generated by Lipschitz perturbations of non-densely defined operators, Differ. Integral Equ., 3 (1990), 1035-1066. |
| [24] | R. A. Horn and C. R. Johnson, Matrix analysis, Cambridge Univ. Press, Cambridge, 1985. |
| [25] | J. K. Hale, Asymptotic Behavior of Dissipative Systems, Math. Surveys Monogr. 25, AMS, Providence, RI, 1988. |
| [26] | C. C. McCluskey, Global stability for an SEI epidemiological model with continuous age-structure in the exposed and infectious classes, Math. Biosci. Eng., 9 (2012), 819-841. |
| [27] | R. Adams and J. Fournier, Sobolev Spaces, Second edition, Pure Appl. Math. 140, Elsevier/Academic Press, Amsterdam, 2003. |
| [28] | P. Magal and X. Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM J. Math. Anal., 37 (2005), 251-275. |
| [29] | W. Wang and X. Q. Zhao, An endemic model in a patchy environment, Math. Biosci., 190 (2004), 97-112. |
| [30] | H. R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM J. Appl. Math., 70 (2009), 188-211. |
| [31] | M. S. Ciupe, B. L. Bivort, D. M. Bortz, et al., Estimating kinetic parameters from HIV primary infection data through the eyes of three different mathematical models, Math. Biosci., 200 (2006), 1-27. |
| [32] | M. Stafford, L. Corey, E. Daar, et al., Modeling plasma virus concentration during primary infection, J. Theor. Biol., 203 (2000), 285-301. |
| [33] | Y. Yang, L. Zou and S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183-191. |
| [34] | G. F. Webb, Theory of nonlinear age-dependent population dynamics, Marcel Dekker, New York, 1985. |
| [35] | C. Castillo-Chavez and H. Thieme, Asymptotically autonomous epidemic models, in Mathematical Population Dynamics: Analysis of Heterogenity (eds. O. Arino, D. Axelrod, M. Kimmel and M. Langlais), Wuerz, (1995), 33-50. |
| [36] | H. R. Thieme, Convergence results and a Poincaré-Bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755-763. |
| [37] | A. Berman and R. J. Plemmons, Nonnegative Matrices in the Mathematical Sciences, Classics in Applied Mathematics 9, SIAM, Philadelphia, 1994. |
| [38] | J. K. Hale and P. Waltman, Persistence in infinite dimensional systems, SIAM J. Math. Anal., 20 (1989), 388-395. |
| [39] | M. Fiedler, Special matrices and their applications in numerical mathematics, Martinus Nijhoff Publishers, The Hague, 1986. |
| [40] | Y. Nakata and G. Röst, Global analysis for spread of infectious diseases via transportation networks, J. Math. Biol., 70 (2015), 1411-1456. |
| [41] | A. T. Raftery, Applied basic science for basic surgical training, Churchill Livingstone, London, 2008. |
| [42] | J. Wang, R. Zhang and T. Kuniya, Global dynamics for a class of age-infection HIV models with nonlinear infection rate, J. Math. Anal. Appl., 432 (2015), 289-313. |
| [43] | C. A. Beauchemin, T. Miura and S. Iwami, Duration of SHIV production by infected cells is not exponentially distributed: Implications for estimates of infection parameters and antiviral efficacy, Sci. Rep., 7 (2017), 42765. |
| [44] | M. Boullé, T. G. Müller, S. Dähling, et al., HIV cell-to-cell spread results in earlier onset of viral gene expression by multiple infections per cell, PLoS Pathog., 12 (2016), e1005964. |
| [45] | K. M. Law, N. L. Komarova, A. W. Yewdall, et al., In vivo HIV-1 cell-to-cell transmission promotes multicopy micro-compartmentalized infection, Cell Rep., 15 (2016), 2771-2783. |
| [46] | A. Sigal, J. T. Kim, A. B. Balazs, et al., Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95. |
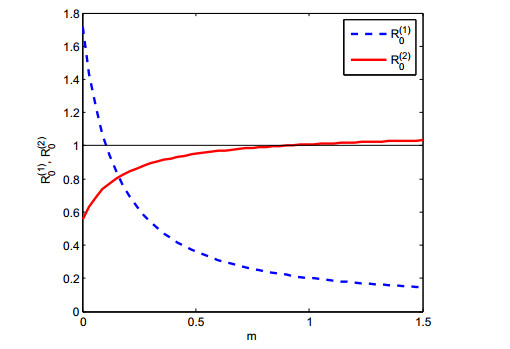
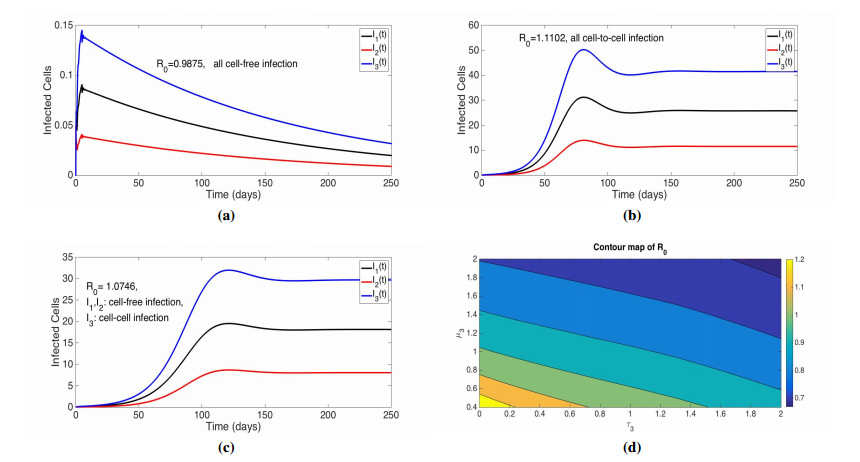
Figures(6)

Cameron J. Browne, Chang-Yuan Cheng. Age-structured viral dynamics in a host with multiple compartments[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 538-574. doi: 10.3934/mbe.2020029







 DownLoad:
DownLoad: