Citation: Huqing Wang, Feng Xiang, Wenbing Zhao, Zhixin Sun. ONS resolution prediction based on Rasch model[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6683-6695. doi: 10.3934/mbe.2019333

| [1] | L. Atzori, A. Iera, G. Morabito, The internet of things: A survey, Comput. Netw., 54(2010), 2787–2805. |
| [2] | C. Chang and C. Li, Algebraic secret sharing using privacy homomorphisms for IoT-based healthcare systems, Math. Biosci. Eng., 16(2019), 3367–3381. |
| [3] | C. Stergiou, K. E. Psannis, A. P. Plageras, et al., Algorithms for efficient digital media transmission over IoT and cloud networking, J. Multi. Inf. Syst., 5(2018), 27–34. |
| [4] | F. X. Yu, I-Japan strategy 2015, Zhong guo Xinxihua, 13(2014), 13–23. |
| [5] | F. Gao, Z. Y. Zhao, K. X. Zhao, et al., Interpretation of the medium-term and long-term strategy of Korean ICT R&D (2013–2017), Sci. Tech. Mgt. Res., 6(2015), 30–34. |
| [6] | W. Ejaz, M. Naeem, A. Shahid, et al., Efficient energy management for the internet of thins in smart cities, IEEE Commun. Mag., 55(2017), 84–91. |
| [7] | C. M. Medaglia and A. Serbanati, An overview of privacy and security issues in the Internet of Things, Proceedings of the 20th Tyrrhenian Workshop on Digital Communications (Italy), Springer, (2010), 389–394. |
| [8] | Y. L. Liu, P. Hua and J. Wang, Verifiable diversity ranking search over encrypted outsourced data, CMC-Comput. Mater. Con., 55(2018), 037–057. |
| [9] | J. Wang, C. W. Ju, Y. Gao, et al., A PSO based energy efficient coverage control algorithm for wireless sensor networks, CMC-Comput. Mater. Con., 56(2018), 433–446. |
| [10] | Z. W. Li and Y. Xie, A security query protocol of ONS in EPC system, 2012 International Conference on Anti-Counterfeiting, Security and Identification (Taipei), IEEE, (2012), Article number: 6325293. |
| [11] | L. J. Zhang and Z. Q. Wu, A controllable trusted and anonymous query mechanism of Internet of Things, Comput. Technol. Develop., 23(2013), 122–125. |
| [12] | C. Fachkha, H. B. Elias and M. Debbabi, Fingerprinting internet DNS amplification DDoS Activities, 2014 6th International Conference on New Technologies, Mobility and Security ( Dubai), OALib J., (2014), article number: 6814019. |
| [13] | A. Friedlander, A. Mankin, W. D. Maughan, et al., DNSSEC: A protocol toward securing the Internet infrastructure, Commu. ACM, 50(2007), 44–50. |
| [14] | M. Dempsky, DNSCurve: Link-Level Security for the Domain Name System, Int. Draft, 2004. |
| [15] | Z. Wu, Y. Zhou and J. Ma, A security transmission model for Internet of Things, Chinese J. Comput., 34(2011), 1351–1364. |
| [16] | W. Ren, L. Ma and Y. Ren, APP: An Ultralightweight scheme to authenticate ONS and protect epc privacy without cryptography in EPCglobal networks, Int J. Distrib. Sens. N., (2013), Article ID: 784618. |
| [17] | P. Danielis, V. Altmann, J. Skodzik, et al., P-DONAS: A P2P-based domain name system in access networks, ACM T. Int. Techn., 15(2015), 1–11. |
| [18] | E. Zhang, X. T. Duan, S. M. Xiu, et al., Server-aided multi-secret sharing scheme for weak computational devices, CMC-Comput. Mater. Con., 56(2018), 401–414. |
| [19] | R. Georg, Probalilistic models for some intelligence and attainment tests, Copenhagen: Institute of Education Research, 1960. |
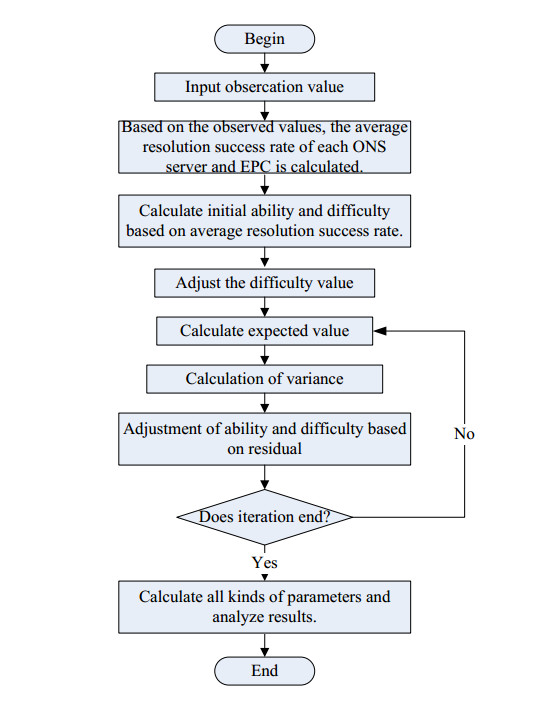
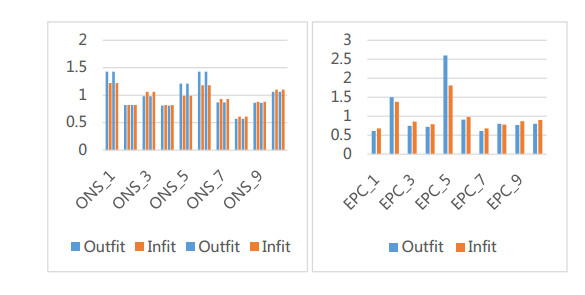
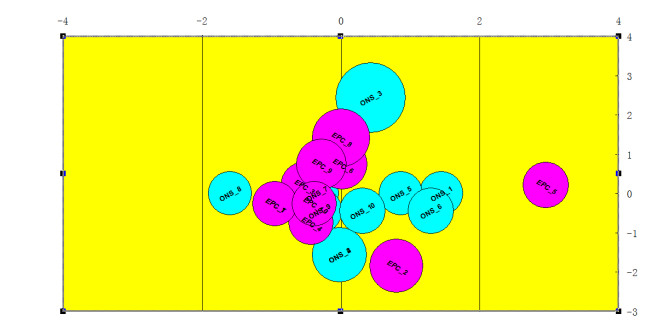
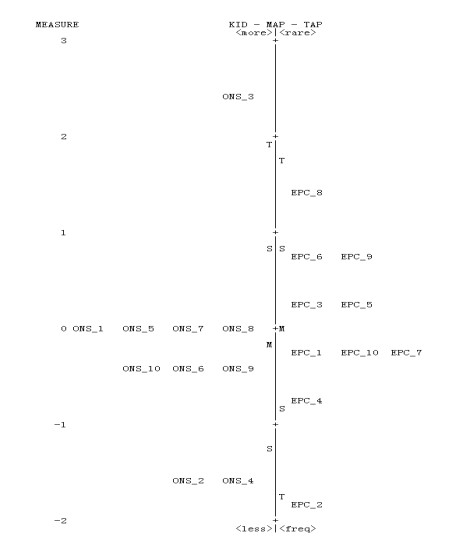
Figures(5) / Tables(2)

Huqing Wang, Feng Xiang, Wenbing Zhao, Zhixin Sun. ONS resolution prediction based on Rasch model[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6683-6695. doi: 10.3934/mbe.2019333







 DownLoad:
DownLoad: