Citation: Carlos Ramos. Animal movement: symbolic dynamics and topological classification[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5464-5489. doi: 10.3934/mbe.2019272

| [1] | P. E. Smouse, S. Focardi, P. R. Moorcroft, et al., Stochastic modelling of animal movement, Phil. Trans. R. Soc. B, 365 (2010), 2201–2211. |
| [2] | R. Nathan, M. Getz, E. Revilla, et al., A movement ecology paradigm for unifying organismal movement research, Proc. Natl Acad. Sci. USA, 105 (2008), 19052–19059. |
| [3] | W. Tang and D. A. Bennett, Agent-based modeling of animal movement: a review, Geography Compass, 4 (2010), 682–700. |
| [4] | I. D. Jonsen, J. M. Flemming and R. A. Myers, Robust state–space modeling of animal movement data, Ecology, 86 (2005), 2874–2880. |
| [5] | T. A. Patterson, L. Thomas, C. Wilcox, et al., State–space models of individual animal movement, Trends Ecol. Evol., 23 (2008), 87–94. |
| [6] | B. T. McClintock, R. King, L. Thomas, et al., A general discrete-time modeling framework for animal movement using multistate random walks, Ecol. Monogr., 82 (2012), 335–349. |
| [7] | R. Langrock, R. King, J Matthiopoulos, et al., Flexible and practical modeling of animal telemetry data: hidden Markov models and extensions, Ecology, 93 (2012), 2336–2342. |
| [8] | S. Petrovskii, A. Mashanova and V. Jansen, Variation in individual walking behavior creates the impression of a Lévy flight, PNAS, 108 (2011), 8704–8707. |
| [9] | P. Tilles, S. Petrovskii and P. Natti, A random walk description of individual animal movement accounting for periods of rest, R. Soc. Open Sci., 3 (2016), 28018645. |
| [10] | J. P. Lampreia and J. Sousa Ramos, Symbolic dynamics of bimodal maps, Portugal. Math., 54 (1997), 1–18. |
| [11] | J. P. Lampreia, R. Severino and J. Sousa Ramos, Renormalizations for cubic maps, Ann. Math. Silesianae, 13 (1999), 257–270. |
| [12] | J. P. Lampreia, R. Severino and J Sousa Ramos, Irreducible complexity of iterated symmetrical bimodal maps, Discrete Dyn. Nat. Soc., 1 (2005), 69–85. |
| [13] | J. Milnor and C. Tresser, On entropy and monotonicity of real cubic maps, Stony Brook, IMS 1998/9 (1998), arXiv:math/9809096. |
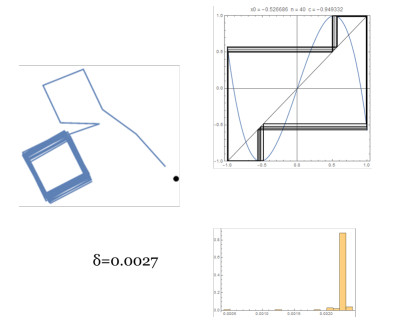
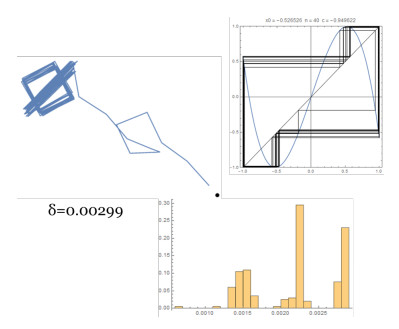
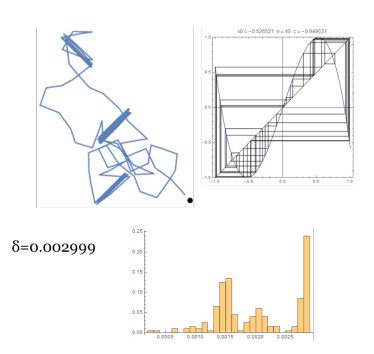
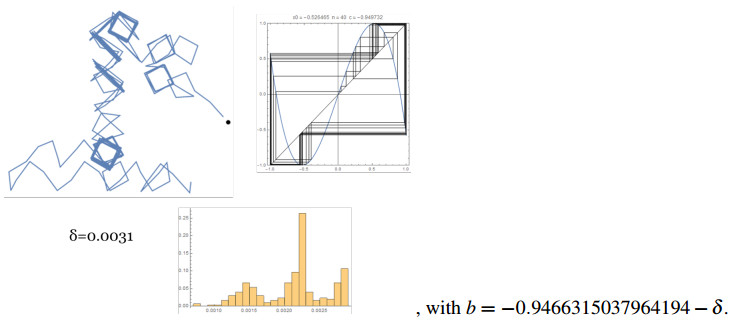

Figures(14)

Carlos Ramos. Animal movement: symbolic dynamics and topological classification[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5464-5489. doi: 10.3934/mbe.2019272







 DownLoad:
DownLoad: