Citation: Thomas Hillen, Kevin J. Painter, Amanda C. Swan, Albert D. Murtha. Moments of von mises and fisher distributions and applications[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 673-694. doi: 10.3934/mbe.2017038

| [1] | [ wikipedia. com, https://en.wikipedia.org/wiki/VonMises%E2%80%93Fisher_distribution, last accessed on 11/22/2016. |
| [2] | [ A. Banerjee,I. S. Dhillon,J. Ghosh,S. Sra, Clustering on the unit hypersphere using von Mises-Fisher distributions, Journal of Machine Learning Research, 6 (2005): 1345-1382. |
| [3] | [ E. Batschelet, null, Circular Statistics in Biology, Academic Press, London, 1981. |
| [4] | [ C. Beaulieu, The basis of anisotropic water diffusion in the nervous system -a technical review, NMR Biomed., 15 (2002): 435-455. |
| [5] | [ R. Bleck, An oceanic general circulation model framed in hybrid isopycnic-cartesian coordinates, Ocean Mod., 4 (2002): 55-88. |
| [6] | [ E. A. Codling,N. Bearon,G. J. Thorn, Thorn, Diffusion about the mean drift location in a biased random walk, Ecology, 91 (2010): 3106-3113. |
| [7] | [ E. A. Codling,M. J. Plank,S. Benhamou, Random walk models in biology, J. Roy. Soc. Interface, 5 (2008): 813-834. |
| [8] | [ C. Darwin, Perception in the lower animals, Nature, 7 (1873): 360. |
| [9] | [ C. Engwer,T. Hillen,M. Knappitsch,C. Surulescu, Glioma follow white matter tracts: A multiscale DTI-based model, J. Math. Biol., 71 (2015): 551-582. |
| [10] | [ N. I. Fisher, null, Statistical Analysis of Circular Data, Cambridge University Press, Cambridge, 1993. |
| [11] | [ A. Giese,L. Kluwe,B. Laube,H. Meissner,M. E. Berens,M. Westphal, Migration of human glioma cells on myelin, Neurosurgery, 38 (1996): 755-764. |
| [12] | [ P. G. Gritsenko,O. Ilina,P. Friedl, Interstitial guidance of cancer invasion, Journal or Pathology, 226 (2012): 185-199. |
| [13] | [ H. Hatzikirou,A. Deutsch,C. Schaller,M. Simaon,K. Swanson, Mathematical modelling of glioblastoma tumour development: A review, Math. Models Meth. Appl. Sci., 15 (2005): 1779-1794. |
| [14] | [ T. Hillen, ${M}^5$ mesoscopic and macroscopic models for mesenchymal motion, J. Math. Biol., 53 (2006): 585-616. |
| [15] | [ T. Hillen, E. Leonard and H. van Roessel, Partial Differential Equations; Theory and Completely Solved Problems, Wiley, Hoboken, NJ, 2012. |
| [16] | [ T. Hillen and K. Painter, Transport and anisotropic diffusion models for movement in oriented habitats, In: Lewis, M., Maini, P., Petrovskii, S. (Eds.), Dispersal, Individual Movement and Spatial Ecology: A Mathematical Perspective, Springer, Heidelberg, 2071 (2013), 177-222. |
| [17] | [ T. Hillen, On the $L^2$-moment closure of transport equations: The general case, Discr.Cont.Dyn.Systems, Series B, 5 (2005): 299-318. |
| [18] | [ A. R. C. James,A. K. Stuart-Smith, Distribution of caribou and wolves in relation to linear corridors, The Journal of Wildlife Management, 64 (2000): 154-159. |
| [19] | [ A. Jbabdi,E. Mandonnet,H. Duffau,L. Capelle,K. Swanson,M. Pelegrini-Issac,R. Guillevin,H. Benali, Simulation of anisotropic growth of low-grade gliomas using diffusion tensor imaging, Mang. Res. Med., 54 (2005): 616-624. |
| [20] | [ J. Kent, The Fisher-Bingham Distribution on the Sphere, J. Royal. Stat. Soc., 1982. |
| [21] | [ E. Konukoglu,O. Clatz,P. Bondiau,H. Delignette,N. Ayache, Extrapolation glioma invasion margin in brain magnetic resonance images: Suggesting new irradiation margins, Medical Image Analysis, 14 (2010): 111-125. |
| [22] | [ K. V. Mardia and P. E. Jupp, Directional Statistics, Wiley, New York, 2000. |
| [23] | [ H. McKenzie,E. Merrill,R. Spiteri,M. Lewis, How linear features alter predator movement and the functional response, Royal Society Interface, 2 (2012): 205-216. |
| [24] | [ P. Moorcroft,M. A. Lewis, null, Mechanistic Home Range Analysis, Princeton University Press, USA, 2006. |
| [25] | [ J. A. Mortimer,A. Carr, Reproduction and migrations of the Ascension Island green turtle (chelonia mydas), Copeia, 1987 (1987): 103-113. |
| [26] | [ P. Mosayebi,D. Cobzas,A. Murtha,M. Jagersand, Tumor invasion margin on the Riemannian space of brain fibers, Medical Image Analysis, 16 (2011): 361-373. |
| [27] | [ A. Okubo and S. Levin, Diffusion and Ecological Problems: Modern Perspectives, Second edition. Interdisciplinary Applied Mathematics, 14. Springer-Verlag, New York, 2001. |
| [28] | [ H. Othmer,S. Dunbar,W. Alt, Models of dispersal in biological systems, Journal of Mathematical Biology, 26 (1988): 263-298. |
| [29] | [ K. Painter, Modelling migration strategies in the extracellular matrix, J. Math. Biol., 58 (2009): 511-543. |
| [30] | [ K. Painter,T. Hillen, Mathematical modelling of glioma growth: The use of diffusion tensor imaging DTI data to predict the anisotropic pathways of cancer invasion, J. Theor. Biol., 323 (2013): 25-39. |
| [31] | [ K. Painter,T. Hillen, Navigating the flow: Individual and continuum models for homing in flowing environments, Royal Society Interface, 12 (2015): 20150,647. |
| [32] | [ K. J. Painter, Multiscale models for movement in oriented environments and their application to hilltopping in butterflies, Theor. Ecol., 7 (2014): 53-75. |
| [33] | [ J. Rao, Molecular mechanisms of glioma invasiveness: The role of proteases, Nature Reviews Cancer, 3 (2003): 489-501. |
| [34] | [ A. Swan, T. Hillen, J. Bowman and A. Murtha, A patient-specific anisotropic diffusion model for brain tumor spread, Bull. Math. Biol., to appear (2017). |
| [35] | [ K. Swanson,E. A. Jr,J. Murray, A quantitative model for differential motility of gliomas in grey and white matter, Cell Proliferation, 33 (2000): 317-329. |
| [36] | [ K. Swanson,R. Rostomily,E. Alvord Jr, Predicting survival of patients with glioblastoma by combining a mathematical model and pre-operative MR imaging characteristics: A proof of principle, British Journal of Cancer, 98 (2008): 113-119. |
| [37] | [ P. Turchin, Quantitative Analysis of Movement, Sinauer Assoc., Sunderland, 1998. |
| [38] | [ C. Xue,H. Othmer, Multiscale models of taxis-driven patterning in bacterial populations, SIAM Journal for Applied Mathematics, 70 (2009): 133-167. |
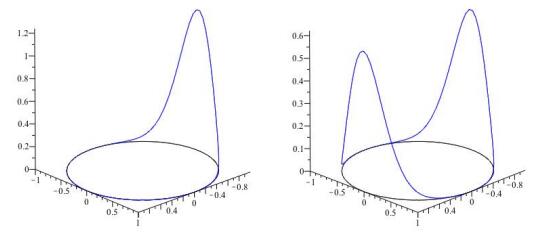
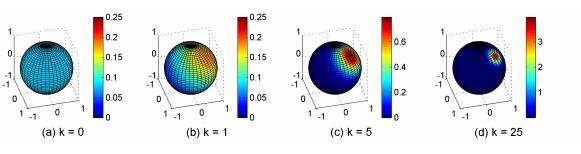
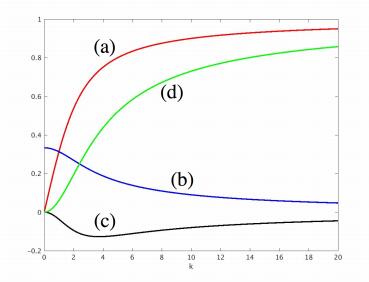
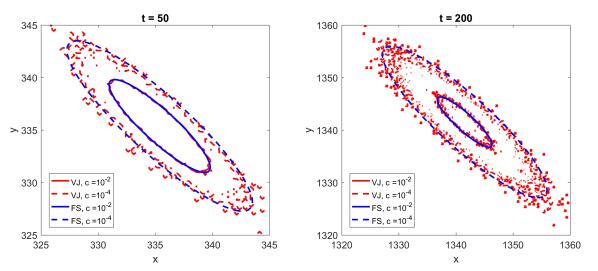


Figures(8)

Thomas Hillen, Kevin J. Painter, Amanda C. Swan, Albert D. Murtha. Moments of von mises and fisher distributions and applications[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 673-694. doi: 10.3934/mbe.2017038







 DownLoad:
DownLoad: