Citation: Thanh Nam Nguyen, Jean Clairambault, Thierry Jaffredo, Benoît Perthame, Delphine Salort. Adaptive dynamics of hematopoietic stem cells and their supporting stroma: a model and mathematical analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4818-4845. doi: 10.3934/mbe.2019243

| [1] | M. C. Mackey, Unified hypothesis for the origin of aplastic anemia and periodic hematopoiesis, Blood, 51 (1978), 941–956. |
| [2] | F. Burns and I. Tannock, On the existence of a G 0 -phase in the cell cycle, Cell Prolif., 3 (1970), 321–334. |
| [3] | L. Lajtha, On DNA labeling in the study of the dynamics of bone marrow cell populations, F. StohlmanJr.(Ed.), The KineticsofCellularProliferation, Gruneand Stratton, NewYork, 173–182. |
| [4] | M. Adimy, S. Bernard, J. Clairambault, et al., Modélisation de la dynamique de l'hématopoïèse normale et pathologique, Hématologie, 14 (2008), 339–350. |
| [5] | M. C. Mackey, C. Ou, L. Pujo-Menjouet, et al., Periodic oscillations of blood cell populations in chronic myelogenous leukemia, SIAM J. Math. Anal., 38 (2006), 166–187. |
| [6] | M. Adimy, F. Crauste and S. Ruan, A mathematical study of the hematopoiesis process with applications to chronic myelogenous leukemia, SIAM J. Appl. Math., 65 (2005), 1328–1352. |
| [7] | L. Pujo-Menjouet, S. Bernard and M. C. Mackey, Long period oscillations in a G 0 model of hematopoietic stem cells, SIAM J. Appl. Dyn. Syst., 4 (2005), 312–332. |
| [8] | M. Adimy, F. Crauste and S. Ruan, Periodic oscillations in leukopoiesis models with two delays, J. Theor. Biol., 242 (2006), 288–299. |
| [9] | A. Ducrot and V. Volpert, On a model of Leukemia development with a spatial cell distribution, Math. Model. Nat. Phenom., 2 (2007), 101–120. |
| [10] | G. Clapp and D. Levy, A review of mathematical models for leukemia and lymphoma, Drug Discov. Today Dis. Models, 16 (2015), 1–6. |
| [11] | L. Pujo-Menjouet, Blood cell dynamics: half of a century of modelling, Math. Model. Nat. Phenom., 11 (2016), 92–115. |
| [12] | B. A. Anthony and D. C. Link, Regulation of hematopoietic stem cells by bone marrow stromal cells, Trends Immunol., 35 (2014), 32–37. |
| [13] | G. Stik, L. Petit, P. Charbord, et al., Vésicules extracellulaires stromales et régulation des cellules souches et progéniteurs hématopoï étiques, médecine/sciences, 34 (2018), 114–116. |
| [14] | P. Charbord, T. Jaffredo and C. Durand, Le cœur moléculaire de la fonction de la niche des cellules souches hématopoï étiques, médecine/sciences, 31 (2015), 12–14. |
| [15] | P. Hirsch, Y. Zhang, R. Tang, et al., Genetic hierarchy and temporal variegation in the clonal history of acute myeloid leukaemia, Nat. commun., 7 (2016), 12475. |
| [16] | P.-E. Jabin and G. Raoul, On selection dynamics for competitive interactions, J. Math. Biol., 63 (2011), 493–517. |
| [17] | C. Pouchol, J. Clairambault, A. Lorz, et al., Asymptotic analysis and optimal control of an integro- differential system modelling healthy and cancer cells exposed to chemotherapy, J. Math. Pures Appl., 116 (2018), 268–308. |
| [18] | C. Pouchol and E. Trélat, Global stability with selection in integro-differential Lotka-Volterra systems modelling trait-structured populations, J. Biol.l Dyn., 12 (2018), 872–893. |
| [19] | G. Barles and B. Perthame, Dirac concentrations in Lotka-Volterra parabolic PDEs, Indiana Univ. Math. J., 57 (2008), 3275–3301. |
| [20] | O. Diekmann, A beginner's guide to adaptive dynamics, Banach Center Publications, 63 (2003), 47–86. |
| [21] | O. Diekmann, P.-E. Jabin, S. Mischler, et al., The dynamics of adaptation: An illuminating example and a hamilton–jacobi approach, Theor. Popul. Biol., 67 (2005), 257–271. |
| [22] | W. Djema, C. Bonnet, F. Mazenc, et al., Control in dormancy or eradication of cancer stem cells: Mathematical modeling and stability issues, J. Theor. Biol., 449 (2018), 103–123. |
| [23] | A. Lorz, S. Mirrahimi and B. Perthame, Dirac mass dynamics in multidimensional nonlocal parabolic equations, Commun Part. Diff. Eq., 36 (2011), 1071–1098. |
| [24] | A. Lorz and B. Perthame, Long-term behaviour of phenotypically structured models, Proc. R. Soc. Lond. Ser. A Math. Phys. Eng. Sci., 470 (2014), 20140089, 10. |
| [25] | G. Meszéna, I. Czibula and S. Geritz, Adaptive dynamics in a 2-patch environment: A toy model for allopatric and parapatric speciation, J. Biol. Syst., 5 (1997), 265–284. |
| [26] | S. Mirrahimi and J.-M. Roquejoffre, A class of Hamilton-Jacobi equations with constraint: uniqueness and constructive approach, J. Differ. Equations, 260 (2016), 4717–4738. |
| [27] | J. D. Murray, Mathematical biology. I, vol. 17 of Interdisciplinary Applied Mathematics, 3rd edition, Springer-Verlag, New York, 2002, An introduction. |
| [28] | B. Perthame, Transport equations in biology, Frontiers in Mathematics, Birkhäuser Verlag, Basel, 2007. |
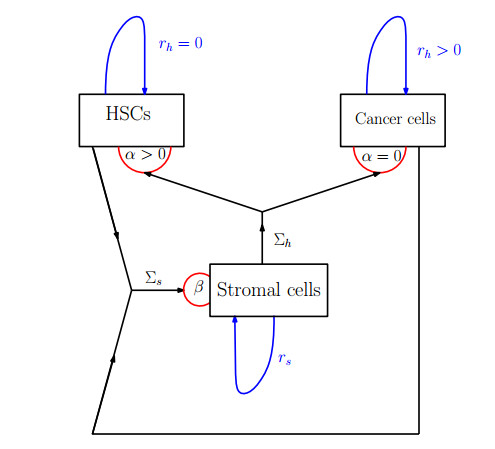
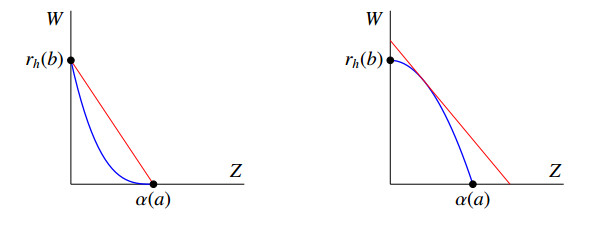
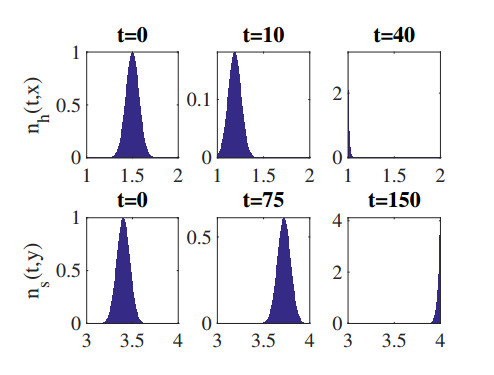
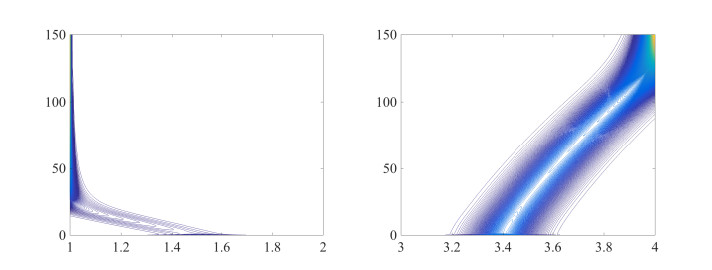
Figures(8) / Tables(1)

Thanh Nam Nguyen, Jean Clairambault, Thierry Jaffredo, Benoît Perthame, Delphine Salort. Adaptive dynamics of hematopoietic stem cells and their supporting stroma: a model and mathematical analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4818-4845. doi: 10.3934/mbe.2019243







 DownLoad:
DownLoad: