Citation: Kamaldeen Okuneye, Ahmed Abdelrazec, Abba B. Gumel. Mathematical analysis of a weather-driven model for the population ecology of mosquitoes[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 57-93. doi: 10.3934/mbe.2018003

| [1] | [ A. Abdelrazec,A. B. Gumel, Mathematical assessment of the role of temperature and rainfall on mosquito population dynamics, Journal of Mathematical Biology, 74 (2017): 1351-1395. |
| [2] | [ F. B. Agusto,A. B. Gumel,P. E. Parham, Qualitative assessment of the role of temperature variations on malaria transmission dynamics, Journal of Biological Systems, 23 (2015): 597-630. |
| [3] | [ N. Ali,K. Marjan,A. Kausar, Study on mosquitoes of Swat Ranizai sub division of Malakand, Pakistan Journal of Zoology, 45 (2013): 503-510. |
| [4] | [ Anopheles Mosquitoes, Centers for Disease Control and Prevention, http://www.cdc.gov/malaria/about/biology/mosquitoes/. Accessed: May, 2016. |
| [5] | [ N. Bacaër, Periodic matrix population models: Growth rate, basic reproduction number and entropy, Bulletin of Mathematical Biology, 71 (2009): 1781-1792. |
| [6] | [ N. Bacaër, Approximation of the basic reproduction number $R_0$ for vector-borne diseases with a periodic vector population, Bulletin of Mathematical Biology, 69 (2007): 1067-1091. |
| [7] | [ N. Bacaër,S. Guernaoui, The epidemic threshold of vector-borne diseases with seasonality, Journal of Mathematical Biology, 53 (2006): 421-436. |
| [8] | [ N. Bacaër,R. Ouifki, Growth rate and basic reproduction number for population models with a simple periodic factor, Mathematical Biosciences, 210 (2007): 647-658. |
| [9] | [ N. Bacaër,X. Abdurahman, Resonance of the epidemic threshold in a periodic environment, Journal of Mathematical Biology, 57 (2008): 649-673. |
| [10] | [ N. Bacaër,H. Ait Dads el, Genealogy with seasonality, the basic reproduction number, and the influenza pandemic, Journal of Mathematical Biology, 62 (2011): 741-762. |
| [11] | [ M. Belda,E. Holtanová,T. Halenka,J. Kalvová, Climate classification revisited: From Köppen to Trewartha, Climate Research, 59 (2014): 1-13. |
| [12] | [ K. Berkelhamer,T. J. Bradley, Mosquito larval development in container habitats: The role of rotting Scirpus californicus, Journal of the American Mosquito Control Association, 5 (1989): 258-260. |
| [13] | [ B. Gates, Gatesnotes: Mosquito Week The Deadliest Animal in the World, https://www.gatesnotes.com/Health/Most-Lethal-Animal-Mosquito-Week. Accessed: May, 2016. |
| [14] | [ S. M. Blower,H. Dowlatabadi, Sensitivity and uncertainty analysis of complex models of disease transmission: An HIV model, as an example, International Statistical Review, 2 (1994): 229-243. |
| [15] | [ P. Cailly,A. Tranc,T. Balenghiene,C. Totyg,P. Ezannoa, A climate-driven abundance model to assess mosquito control strategies, Ecological Modelling, 227 (2012): 7-17. |
| [16] | [ J. Cariboni,D. Gatelli,R. Liska,A. Saltelli, A. The role of sensitivity analysis in ecological modeling, Ecological Modeling, 203 (2007): 167-182. |
| [17] | [ J. Carr, null, Applications of Centre Manifold Theory, , Springer-Verlag, New York, 1981. |
| [18] | [ C. Castillo-Chavez,B. Song, Dynamical models of tuberculosis and their applications, Mathematical Bioscience Engineering, 1 (2004): 361-404. |
| [19] | [ N. Chitnis,J. M. Cushing,J. M. Hyman, Bifurcation analysis of a mathematical model for malaria transmission, SIAM Journal on Applied Mathematics, 67 (2006): 24-45. |
| [20] | [ S. Chow,C. Li,D. Wang, null, Normal Forms and Bifurcation of Planar Vector Fields, , Cambridge University Press, Cambridge, 1994. |
| [21] | [ J. Couret,E. Dotson,M. Q. Benedict, Temperature, Larval diet, and density effects on development rate and survival of Aedes aegypti (Diptera: Culicidae), PLoS One, 9 (2014). |
| [22] | [ J. M. O. Depinay,C. M. Mbogo,G. Killeen,B. Knols,J. Beier, A simulation model of African Anopheles ecology and population dynamics for the analysis of malaria transmission, Malaria Journal, 3 (2004): p29. |
| [23] | [ O. Diekmann,J. Heesterbeek,J. Metz, On the definition and the computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous populations, Journal of Mathematical Biology, 28 (1990): 365-382. |
| [24] | [ F. Dufois, Assessing inter-annual and seasonal variability Least square fitting with Matlab: Application to SSTs in the vicinity of Cape Town, http://www.eamnet.eu/cms/sites/eamnet.eu/files/Least_square_fitting_with_Matlab-Francois_Dufois.pdf. Accessed: October, 2016. |
| [25] | [ Durban Monthly Climate Average, South Africa, http://www.worldweatheronline.com/Durban-weather-averages/Kwazulu-Natal/ZA.aspx. Accessed: May 2016. |
| [26] | [ J. Dushoff,W. Huang,C. Castillo-Chavez, Backward bifurcations and catastrophe in simple models of fatal diseases, Journal of Mathematical Biology, 36 (1998): 227-248. |
| [27] | [ T. G. George, Positive Definite Matrices and Sylvester's Criterion, The American Mathematical Monthly, 98 (1991): 44-46. |
| [28] | [ H. M. Giles,D. A. Warrel, null, Bruce-Chwatt's Essential Malariology, 3rd edition, Heinemann Medical Books, Portsmouth, NH, 1993. |
| [29] | [ J. E. Gimnig,M. Ombok,S. Otieno,M. G. Kaufman,J. M. Vulule,E. D. Walker, Density-dependent development of Anopheles gambiae (Diptera: Culicidae) larvae in artificial habitats, Journal of Medical Entomology, 39 (2002): 162-172. |
| [30] | [ R. E. Harbach, Mosquito Taxonomic Inventory, (2011). http://mosquito-taxonomic-inventory.info/simpletaxonomy/term/6045. Accessed: May, 2016. |
| [31] | [ D. Hershkowitz, Recent directions in matrix stability, Linear Algebra and its Applications, 171 (1992): 161-186. |
| [32] | [ W. M. Hirsch,H. Hanisch,J. P. Gabriel, Differential equation models for some parasitic infections: Methods for the study of asymptotic behavior, Communications on Pure and Applied Mathematics, 38 (1985): 733-753. |
| [33] | [ S. S. Imbahale,K. P. Paaijmans,W. R. Mukabana,R. van Lammeren,A. K. Githeko,W. Takken, A longitudinal study on Anopheles mosquito larval abundance in distinct geographical and environmental settings in western Kenya, Malaria Journal, 10 (2011). |
| [34] | [ K. C. Kain,J. S. Keystone, Malaria in travelers, Infectious Disease Clinics, 12 (1998): 267-284. |
| [35] | [ V. Kothandaraman, Air-water temperature relationship in Illinois River, Water Resources Bulletin, 8 (1972): 38-45. |
| [36] | [ Lagos Monthly Climate Average, Nigeria, http://www.worldweatheronline.com/lagos-weather-averages/lagos/ng.aspx. Accessed: May 2016. |
| [37] | [ V. Lakshmikantham,S. Leela, null, Differential and Integral Inequalities: Theory and Applications, , Academic Press, New York-London, 1969. |
| [38] | [ V. Laperriere,K. Brugger,F. Rubel, Simulation of the seasonal cycles of bird, equine and human West Nile virus cases, Preventive Veterinary Medicine, 88 (2011): 99-110. |
| [39] | [ J. P. LaSalle, The Stability of Dynamical Systems Regional Conference Series in Applied Mathematics. SIAM Philadephia. 1976. |
| [40] | [ Y. Lou,X.-Q. Zhao, A climate-based malaria transmission model with structured vector population, SIAM Journal on Applied Mathematics, 70 (2010): 2023-2044. |
| [41] | [ A. M. Lutambi,M. A. Penny,T. Smith,N. Chitnis, Mathematical modelling of mosquito dispersal in a heterogeneous environment, Journal of Mathematical Biosciences, 241 (2013): 198-216. |
| [42] | [ P. Magal,X.-Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM Journal on Mathematical Analysis, 37 (2005): 251-275. |
| [43] | [ Malaria Atlas Project: Mosquito Malaria Vectors, http://www.map.ox.ac.uk/explore/mosquito-malaria-vectors/, Accessed: May: 2016. |
| [44] | [ S. Marino,I. B. Hogue,C. J. Ray,D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, Journal of Theoretical Biology, 254 (2008): 178-196. |
| [45] | [ M. D. Mckay,R. J. Beckman,W. J. Conover, Comparison of 3 methods for selecting values of input variables in the analysis of output from a computer code, Technometrics, 21 (1979): 239-245. |
| [46] | [ R. G. McLeod,J. F. Brewster,A. B. Gumel,D. A. Slonowsky, Sensitivity and uncertainty analyses for a SARS model with time-varying inputs and outputs, Mathematical Biosciences and Engineering, 3 (2006): 527-544. |
| [47] | [ E. A. Mordecai, Optimal temperature for malaria transmission is dramatically lower than previously predicted, Ecology Letters, 16 (2013): 22-30. |
| [48] | [ Mosquito Life Cycle. American Mosquito Control Association, http://www.mosquito.org/life-cycle, Accessed: May, 2016. |
| [49] | [ Mosquitoes of Michigan -Their Biology and Control, Michigan Mosquito Control Organization, 2013. http://www.mimosq.org/mosquitobiology/mosquitobiology.htm. Accessed: May: 2015. |
| [50] | [ Nairobi Monthly Climate Average, Kenya, http://www.worldweatheronline.com/nairobi-weather-averages/nairobi-area/ke.aspx. Accessed: May 2016. |
| [51] | [ G. A. Ngwa, On the population dynamics of the malaria vector, Bulletin of Mathematical Biology, 68 (2006): 2161-2189. |
| [52] | [ G. A. Ngwa,A. M. Niger,A. B. Gumel, Mathematical assessment of the role of non-linear birth and maturation delay in the population dynamics of the malaria vector, Applied Mathematics and Computation, 217 (2010): 3286-3313. |
| [53] | [ A. M. Niger,A. B. Gumel, Mathematical analysis of the role of repeated exposure on malaria transmission dynamics, Differential Equations and Dynamical Systems, 16 (2008): 251-287. |
| [54] | [ T. E. Nkya,I. Akhouayri,W. Kisinza,J. P. David, Impact of environment on mosquito response to pyrethroid insecticides: Facts evidences and prospects, Insect Biochemistry and Molecular Biology, 43 (2013): 407-416. |
| [55] | [ K. O. Okuneye,A. B. Gumel, Analysis of a temperature-and rainfall-dependent model for malaria transmission Dynamics, Mathematical Biosciences, 287 (2017): 72-92. |
| [56] | [ H. J. Overgaard,Y. Tsude,W. Suwonkerd,M. Takagi, Characteristics of Anopheles minimus (Diptera: Culicidae) larval habitats in northern Thailand, Environmental Entomology, 31 (2002): 134-141. |
| [57] | [ K. P. Paaijmans,S. S. Imbahale,M. B. Thomas,W. Takken, Relevant microclimate for determining the development rate of malaria mosquitoes and possible implications of climate change, Malaria Journal, 9 (2010): p196. |
| [58] | [ K. P. Paaijmans,M. O. Wandago,A. K. Githeko,W. Takken, Unexpected high losses of Anopheles gambiae larvae due to rainfall, PLOS One, 2 (2007). |
| [59] | [ P. E. Parham,E. Michael, Modeling the effects of weather and climate change on malaria transmission, Environmental Health Perspectives, 118 (2010): 620-626. |
| [60] | [ P. E. Parham,D. Pople,C. Christiansen-Jucht,S. Lindsay,W. Hinsley,E. Michael, Modeling the role of environmental variables on the population dynamics of the malaria vector Anopheles gambiae sensu stricto, Malaria Journal, 11 (2012): p271. |
| [61] | [ P. C. Park, A new proof of Hermite's stability criterion and a generalization of Orlando's formula, International Journal of Control, 26 (2012): 197-206. |
| [62] | [ J. M. Pilgrim, X. Fang and H. G. Stefan, Correlations of Minnesota Stream Water Temperatures with Air Temperatures Project Report 382, prepared for National Agricultural Water Quality Laboratory Agricultural Research Service U. S. Department of Agriculture Durant, Oklahoma, 1995. |
| [63] | [ T. Porphyre,D. J. Bicout,P. Sabatier, Modelling the abundance of mosquito vectors versus flooding dynamics, Ecological Modelling, 183 (2005): 173-181. |
| [64] | [ E. B. Preud'homme and H. G. Stefan, Relationship Between Water Temperatures and Air Temperatures for Central U. S. Streams Project Report No. 333, prepared for Environmental Research Laboratory U. S. Environmental Protection Agency Duluth, Minnesota, 1992. |
| [65] | [ F. Rubel,K. Brugger,M. Hantel,S. Chvala-Mannsberger,T. Bakonyi,H. Weissenbock,N. Nowotny, Explaining Usutu virus dynamics in Austria: Model development and calibration, Preventive Veterinary Medicine, 85 (2008): 166-186. |
| [66] | [ M. A. Safi,M. Imran,A. B. Gumel, Threshold dynamics of a non-autonomous SEIRS model with quarantine and isolation, Theory in Biosciences, 131 (2012): 19-30. |
| [67] | [ J. Shaman,J. Day, Reproductive phase locking of mosquito populations in response to rainfall frequency, Plos One, 2 (2007): p331. |
| [68] | [ O. Sharomi,C. N. Podder,A. B. Gumel,E. H. Elbasha,J. Watmough, Role of incidence function in vaccine-induced backward bifurcation in some HIV models, Mathematical Biosciences, 210 (2007): 436-463. |
| [69] | [ H. L. Smith, null, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, , American Mathematical Society, 1995. |
| [70] | [ H. L. Smith,P. Waltman, Perturbation of a globally stable steady state, American Mathematical Society, 127 (1999): 447-453. |
| [71] | [ H. R. Thieme, Convergence results and a Poincaré-Bendixson trichotomy for asymptotically autonomous differential equations, Journal of Mathematical Biology, 30 (1992): 755-763. |
| [72] | [ H. R. Thieme, Persistence under relaxed point dissipativity (with application to an endemic model), SIAM Journal on Mathematical Analysis, 24 (1993): 407-435. |
| [73] | [ P. Van den Driessche,J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Mathematical Biosciences, 180 (2002): 29-48. |
| [74] | [ E. Van Handel, Nutrient accumulation in three mosquitoes during larval development and its effect on young adults, Journal of the American Mosquito Control Association, 4 (1988): 374-376. |
| [75] | [ W. Wang,X.-Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, Journal of Dynamics and Differential Equations, 20 (2008): 699-717. |
| [76] | [ World Health Organization, A global brief on vector-borne diseases, 2014. |
| [77] | [ World Health Organization, World health report. Executive summary, Insect-borne diseases, 1996. |
| [78] | [ World Health Organization, WHO global health days, http://www.who.int/campaigns/world-health-day/2014/vector-borne-diseases/en/. Accessed: June, 2016. |
| [79] | [ P. Wu,G. Lay,R. Guo,Y. Lin,C. Lung,J. Su, Higher temperature and urbanization affect the spatial patterns of dengue fever transmission in subtropical Taiwan, Science of The Total Environment, 407 (2009): 2224-2233. |
| [80] | [ F. Zhang,X.-Q. Zhao, A periodic epidemic model in a patchy environment, Journal of Mathematical Analysis and Applications, 325 (2007): 496-516. |
| [81] | [ Z. Zhang, T. W. Ding, T. Huang and Z. Dong, Qualitative Theory of Differential Equations American Mathematical, 2006. |
| [82] | [ X.-Q. Zhao, null, Dynamical Systems in Population Biology, , Springer, New York, 2003. |
| [83] | [ X.-Q. Zhao, Permanence implies the existence of interior periodic solutions for FDEs, International Journal of Qualitative Theory of Differential Equations and Applications, 2 (2008): 125-137. |
| [84] | [ X.-Q. Zhao, Uniform persistence and periodic coexistence states in infinite-dimensional periodic semiflows with applications, Canadian Applied Mathematics Quarterly, 3 (1995): 473-495. |
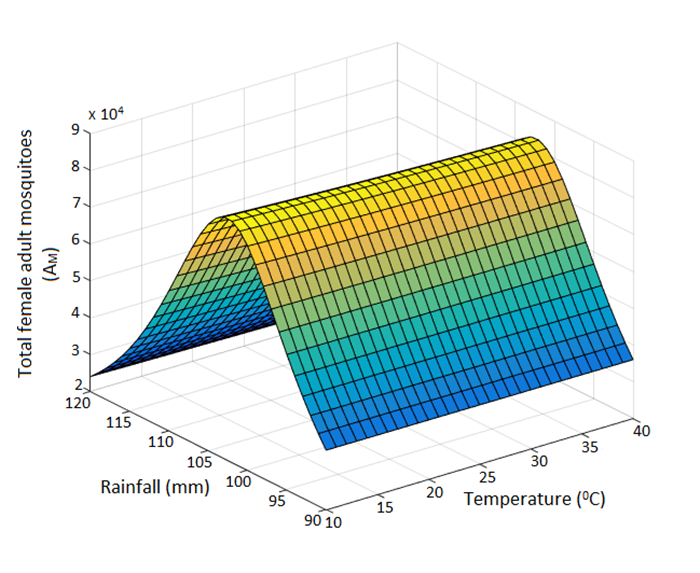
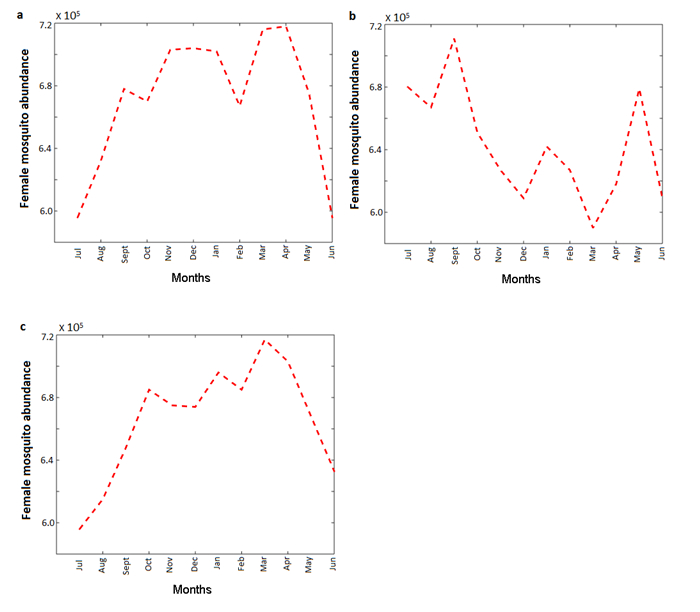
Figures(6) / Tables(9)

Kamaldeen Okuneye, Ahmed Abdelrazec, Abba B. Gumel. Mathematical analysis of a weather-driven model for the population ecology of mosquitoes[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 57-93. doi: 10.3934/mbe.2018003







 DownLoad:
DownLoad: