In general, there is an imperative to amalgamate timely interventions and comprehensive measures for the efficacious control of infectious diseases. The deployment of such measures is intricately tied to the system's state and its transmission rate, presenting formidable challenges for stability and bifurcation analyses. In our pursuit of devising qualitative techniques for infectious disease analysis, we introduced a model that incorporates state-dependent transmission interventions. Through the introduction of state-dependent control, characterized by a non-linear action threshold contingent upon the combination of susceptible population density and its rate of change, we employ analytical methods to scrutinize various facets of the model. This encompasses addressing the existence, stability, and bifurcation phenomena concerning disease-free periodic solutions (DFPS). The analysis of the established Poincaré map leads us to the conclusion that DFPS indeed exists and maintains stability under specific conditions. Significantly, we have formulated a distinctive single-parameter family of discrete mappings, leveraging the bifurcation theorems of discrete maps to dissect the transcritical bifurcations around DFPS with respect to parameters such as $ ET $ and $ \eta_{1} $. Under particular conditions, these phenomena may give rise to effects like backward bifurcation and bistability. Through the analytical methodologies developed in this study, our objective is to unveil a more comprehensive understanding of infectious disease models and their potential relevance across diverse domains.
Citation: Yongfeng Li, Song Huang, Zhongyi Xiang. A state-dependent impulsive system with ratio-dependent action threshold for investigating SIR model[J]. AIMS Mathematics, 2024, 9(2): 4781-4804. doi: 10.3934/math.2024231
In general, there is an imperative to amalgamate timely interventions and comprehensive measures for the efficacious control of infectious diseases. The deployment of such measures is intricately tied to the system's state and its transmission rate, presenting formidable challenges for stability and bifurcation analyses. In our pursuit of devising qualitative techniques for infectious disease analysis, we introduced a model that incorporates state-dependent transmission interventions. Through the introduction of state-dependent control, characterized by a non-linear action threshold contingent upon the combination of susceptible population density and its rate of change, we employ analytical methods to scrutinize various facets of the model. This encompasses addressing the existence, stability, and bifurcation phenomena concerning disease-free periodic solutions (DFPS). The analysis of the established Poincaré map leads us to the conclusion that DFPS indeed exists and maintains stability under specific conditions. Significantly, we have formulated a distinctive single-parameter family of discrete mappings, leveraging the bifurcation theorems of discrete maps to dissect the transcritical bifurcations around DFPS with respect to parameters such as $ ET $ and $ \eta_{1} $. Under particular conditions, these phenomena may give rise to effects like backward bifurcation and bistability. Through the analytical methodologies developed in this study, our objective is to unveil a more comprehensive understanding of infectious disease models and their potential relevance across diverse domains.

| [1] |
O. Diekmann, J. A. P. Heesterbeek, J. A. Metz, On the definition and the computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324

|
| [2] |
A. B. Sabin, Measles, killer of millions in developing countries: strategy for rapid elimination and continuing control, Eur. J. Epidemiol., 7 (1991), 1–22. https://doi.org/10.1007/BF00221337 doi: 10.1007/BF00221337
|
| [3] |
C. A. de Quadros, J. K. Andrus, J. M. Olivé, C. M. da Silveira, R. M. Eikhof, P. Carrasco, et al., Eradication of poliomyelitis: progress in the Americas, Pediatr. Infect. Dis. J., 10 (1991), 222–229. https://doi.org/10.1097/00006454-199103000-00011 doi: 10.1097/00006454-199103000-00011
|
| [4] | M. Ramsay, N. Gay, E. Miller, M. Rush, J. White, P. Morgan-Capner, et al., The epidemiology of measles in England and Wales: rationale for the 1994 national vaccination campaign, Commun. Dis. Rep., 4 (1994), R141–R146. |
| [5] |
A. Khan, J. F. Gómez-Aguilar, T. S. Khan, H. Khan, Stability analysis and numerical solutions of fractional order HIV/AIDS model, Chaos Solitons Fract., 122 (2019), 119–128. https://doi.org/10.1016/j.chaos.2019.03.022 doi: 10.1016/j.chaos.2019.03.022
|
| [6] |
H. Khan, J. F. Gómez-Aguilar, A. Alkhazzan, A. Khan, A fractional order HIV‐TB coinfection model with nonsingular Mittag‐Leffler law, Math. Methods Appl. Sci., 43 (2020), 6. https://doi.org/10.1002/mma.6155 doi: 10.1002/mma.6155
|
| [7] |
A. Khan, H. M. Alshehri, T. Abdeljawad, Q. M. Al-Mdallal, H. Khan, Stability analysis of fractional nabla difference COVID-19 model, Results Phys., 22 (2021), 103888. https://doi.org/10.1016/j.rinp.2021.103888 doi: 10.1016/j.rinp.2021.103888
|
| [8] |
R. Begum, O. Tunç, H. Khan, H. Gulzar, A. Khan, A fractional order Zika virus model with Mittag-Leffler kernel, Chaos Solitons Fract., 146 (2021), 110898. https://doi.org/10.1016/j.chaos.2021.110898 doi: 10.1016/j.chaos.2021.110898
|
| [9] |
M. Aslam, R. Murtaza, T. Abdeljawad, G. U. Rahman, A. Khan, H. Khan, et al., A fractional order HIV/AIDS epidemic model with Mittag-Leffler kernel, Adv. Differ. Equations, 2021 (2021), 107. https://doi.org/10.1186/s13662-021-03264-5 doi: 10.1186/s13662-021-03264-5
|
| [10] |
Q. T. Ain, A. Khan, T. Abdeljawad, J. F. Gómez-Aguilar, S. Riaz, Dynamical study of varicella-zoster virus model in sense of Mittag-Leffler kernel, Int. J. Biomath., 17 (2024), 2350027. https://doi.org/10.1142/S1793524523500274 doi: 10.1142/S1793524523500274
|
| [11] |
G. R. Jiang, Q. G. Yang, Periodic solutions and bifurcation in an SIS epidemic model with birth pulses, Math. Comput. Modell., 50 (2009), 498–508. https://doi.org/10.1016/j.mcm.2009.04.021 doi: 10.1016/j.mcm.2009.04.021
|
| [12] |
Q. Q. Zhang, S. Y. Tang, X. F. Zou, Rich dynamics of a predator-prey system with state-dependent impulsive controls switching between two means, J. Differ. Equations, 364 (2023), 336–377. https://doi.org/10.1016/j.jde.2023.03.030 doi: 10.1016/j.jde.2023.03.030
|
| [13] |
S. J. Gao, L. S. Chen, Z. D. Teng, Impulsive vaccination of an SEIRS model with time delay and varying total population size, Bull. Math. Biol., 69 (2007), 731–745. https://doi.org/10.1007/s11538-006-9149-x doi: 10.1007/s11538-006-9149-x
|
| [14] |
J. C. Panetta, A mathematical model of periodically pulsed chemotherapy: tumor recurrence and metastasis in a competitive environment, Bull. Math. Biol., 58 (1996), 425–447. https://doi.org/10.1007/BF02460591 doi: 10.1007/BF02460591
|
| [15] |
S. Bunimovich-Mendrazitsky, H. Byrne, L. Stone, Mathematical model of pulsed immunotherapy for superficial bladder cancer, Bull. Math. Biol., 70 (2008), 2055–2076. https://doi.org/10.1007/s11538-008-9344-z doi: 10.1007/s11538-008-9344-z
|
| [16] |
Q. Li, Y. N. Xiao, Dynamical behavior and bifurcation analysis of the SIR model with continuous treatment and state-dependent impulsive control, Int. J. Bifurcat. Chaos, 29 (2019), 1950131. https://doi.org/10.1142/S0218127419501311 doi: 10.1142/S0218127419501311
|
| [17] |
Z. L. He, J. G. Li, L. F. Nie, Z. Zhao, Nonlinear state-dependent feedback control strategy in the SIR epidemic model with resource limitation, Adv. Differ. Equations, 2017 (2017), 209. https://doi.org/10.1186/s13662-017-1229-8 doi: 10.1186/s13662-017-1229-8
|
| [18] |
S. Y. Tang, Y. N. Xiao, D. Clancy, New modelling approach concerning integrated disease control and cost-effectivity, Nonlinear Anal., 63 (2005), 439–471. https://doi.org/10.1016/j.na.2005.05.029 doi: 10.1016/j.na.2005.05.029
|
| [19] |
L. F. Nie, Z. D. Teng, A. Torres, Dynamic analysis of an SIR epidemic model with state dependent pulse vaccination, Nonlinear. Anal., 13 (2012), 1621–1629. https://doi.org/10.1016/j.nonrwa.2011.11.019 doi: 10.1016/j.nonrwa.2011.11.019
|
| [20] |
L. F. Nie, Z. D. Teng, B. Z. Guo, A state dependent pulse control strategy for a SIRS epidemic system, Bull. Math. Biol., 75 (2013), 1697–1715. https://doi.org/10.1007/s11538-013-9865-y doi: 10.1007/s11538-013-9865-y
|
| [21] |
X. W. Yu, S. L. Yuan, T. H. Zhang, Asymptotic properties of stochastic nutrient-plankton food chain models with nutrient recycling, Nonlinear. Anal., 34 (2019), 209–225. https://doi.org/10.1016/j.nahs.2019.06.005 doi: 10.1016/j.nahs.2019.06.005
|
| [22] |
D. D. Fang, Y. Z. Pei, Y. F. Lv, L. S. Chen, Periodicity induced by state feedback controls and driven by disparate dynamics of a herbivore-plankton model with cannibalism, Nonlinear Dyn., 90 (2017), 2657–2672. https://doi.org/10.1007/s11071-017-3829-y doi: 10.1007/s11071-017-3829-y
|
| [23] |
D. Z. Li, Y. Liu, H. D. Cheng, Dynamic complexity of a phytoplankton-fish model with the impulsive feedback control by means of poincare map, Complexity, 2020 (2020), 8974763. https://doi.org/10.1155/2020/8974763 doi: 10.1155/2020/8974763
|
| [24] |
J. Yang, S. Y. Tang, Holling type Ⅱ predator-prey model with nonlinear pulse as state-dependent feedback control, J. Comput. Appl. Math., 291 (2016), 225–241. https://doi.org/10.1016/j.cam.2015.01.017 doi: 10.1016/j.cam.2015.01.017
|
| [25] |
Z. Z. Shi, H. D. Cheng, Y. Liu, Y. H. Wang, Optimization of an integrated feedback control for a pest management predator-prey model, Math. Biosci. Eng., 16 (2019), 7963–7981. https://doi.org/10.3934/mbe.2019401 doi: 10.3934/mbe.2019401
|
| [26] |
I. U. Khan, S. Y. Tang, The impulsive model with pest density and its change rate dependent feedback control, Discrete Dyn. Nat. Soc., 2020 (2020), 4561241. https://doi.org/10.1155/2020/4561241 doi: 10.1155/2020/4561241
|
| [27] |
T. T. Li, W. C. Zhao, Periodic solution of a neutral delay leslie predator-prey model and the effect of random perturbation on the smith growth model, Complexity, 2020 (2020), 8428269. https://doi.org/10.1155/2020/8428269 doi: 10.1155/2020/8428269
|
| [28] |
Y. N. Li, Y. Li, Y. Liu, H. D. Cheng, Stability analysis and control optimization of a prey-predator model with linear feedback control, Discrete Dyn. Nat. Soc., 2018 (2018), 4945728. https://doi.org/10.1155/2018/4945728 doi: 10.1155/2018/4945728
|
| [29] |
H. J. Guo, L. S. Chen, X. Y. Song, Qualitative analysis of impulsive state feedback control to an algae-fish system with bistable property, Appl. Math. Comput., 271 (2015), 905–922. https://doi.org/10.1016/j.amc.2015.09.046 doi: 10.1016/j.amc.2015.09.046
|
| [30] |
Q. Q. Zhang, S. Y. Tang, Bifurcation analysis of an ecological model with nonlinear state–dependent feedback control by poincaré map defined in phase set, Commun. Nonlinear Sci., 108 (2022), 106212. https://doi.org/10.1016/j.cnsns.2021.106212 doi: 10.1016/j.cnsns.2021.106212
|
| [31] |
Y. Z. Wu, G. Y. Tang, C. C. Xiang, Dynamic analysis of a predator-prey state-dependent impulsive model with fear effect in which action threshold depending on the prey density and its changing rate, Math. Biosci. Eng., 19 (2022), 13152–13171. https://doi.org/10.3934/mbe.2022615 doi: 10.3934/mbe.2022615
|
| [32] |
Y. Tian, Y. Gao, K. B. Sun, Global dynamics analysis of instantaneous harvest fishery model guided by weighted escapement strategy, Chaos Solitons Fract., 164 (2022), 112597. https://doi.org/10.1016/j.chaos.2022.112597 doi: 10.1016/j.chaos.2022.112597
|
| [33] |
I. U. Khan, S. Y. Tang, B. Tang, The state-dependent impulsive model with action threshold depending on the pest density and its changing rate, Complexity, 2019 (2019), 6509867. https://doi.org/10.1155/2019/6509867 doi: 10.1155/2019/6509867
|
| [34] |
G. Wang, M. Yi, S. Y. Tang, Dynamics of an antitumour model with pulsed radioimmunotherapy, Comput. Math. Methods Med., 2022 (2022), 4692772. https://doi.org/10.1155/2022/4692772 doi: 10.1155/2022/4692772
|
| [35] |
W. Li, T. H. Zhang, Y. F. Wang, H. D. Cheng, Dynamic analysis of a plankton-herbivore state-dependent impulsive model with action threshold depending on the density and its changing rate, Nonlinear Dyn., 107 (2022), 2951–2963. https://doi.org/10.1007/s11071-021-07022-w doi: 10.1007/s11071-021-07022-w
|
| [36] |
B. Tang, W. Q. Zhao, Sliding dynamics and bifurcations of a filippov system with nonlinear threshold control, Int. J. Bifurcat. Chaos, 31 (2021), 2150214. https://doi.org/10.1142/S021812742150214X doi: 10.1142/S021812742150214X
|
| [37] |
Y. F. Li, S. Huang, X. Y. Song, Global dynamic analysis of a nonlinear state-dependent feedback control SIR model with saturation incidence, Eur. Phys. J. Plus, 138 (2023), 636. https://doi.org/10.1140/epjp/s13360-023-04277-7 doi: 10.1140/epjp/s13360-023-04277-7
|
| [38] |
T. Y. Cheng, S. Y. Tang, R. A. Cheke, Threshold dynamics and bifurcation of a state-dependent feedback nonlinear control susceptible-infected-recovered model, J. Comput. Nonlinear Dyn., 14 (2019), 071001. https://doi.org/10.1115/1.4043001 doi: 10.1115/1.4043001
|
| [39] |
Q. Q. Zhang, B. Tang, S. Y. Tang, Vaccination threshold size and backward bifurcation of SIR model with state-dependent pulse control, J. Theor. Biol., 455 (2018), 75–85. https://doi.org/10.1016/j.jtbi.2018.07.010 doi: 10.1016/j.jtbi.2018.07.010
|
| [40] |
R. J. Smith, E. J. Schwartz, Predicting the potential impact of a cytotoxic T-lymphocyte HIV vaccine: How often should you vaccinate and how strong should the vaccine be? Math. Biosci., 212 (2008), 180–187. https://doi.org/10.1016/j.mbs.2008.02.001 doi: 10.1016/j.mbs.2008.02.001
|
| [41] |
B. Tang, Y. N. Xiao, S. Y. Tang, R. A. Cheke, A feedback control model of comprehensive therapy for treating immunogenic tumours, Int. J. Bifurcat. Chaos, 26 (2016), 1650039. https://doi.org/10.1142/S0218127416500395 doi: 10.1142/S0218127416500395
|
| [42] |
Z. Q. Liang, G. P. Pang, X. P. Zeng, Y. H. Liang, Qualitative analysis of a predator-prey system with mutual interference and impulsive state feedback control, Nonlinear Dyn., 87 (2017), 1495–1509. https://doi.org/10.1007/s11071-016-3129-y doi: 10.1007/s11071-016-3129-y
|
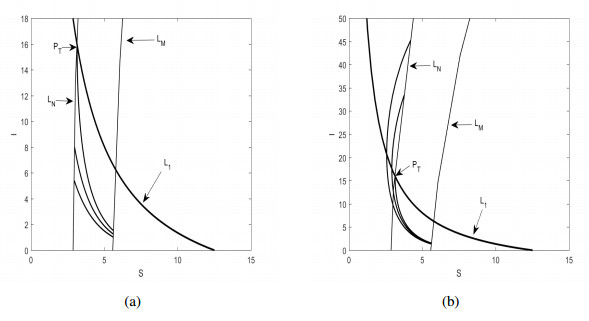
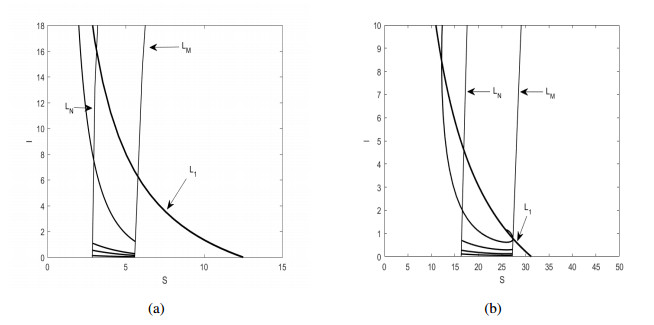
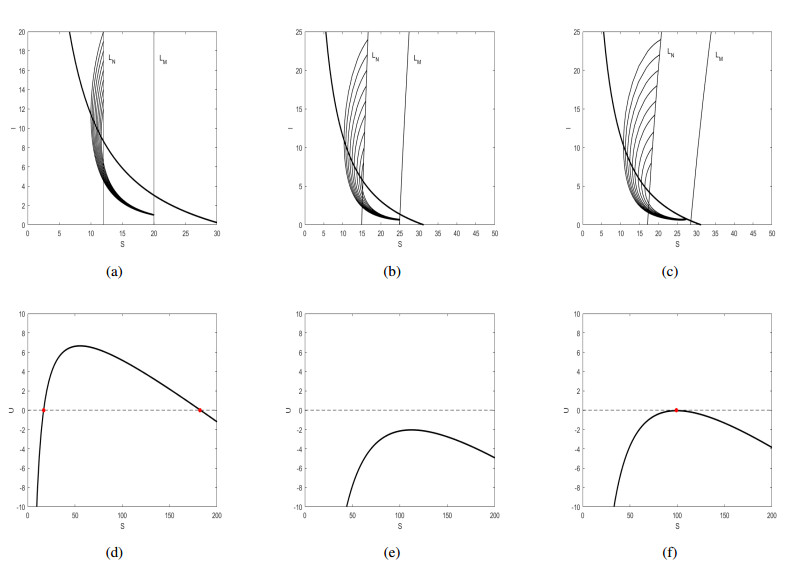
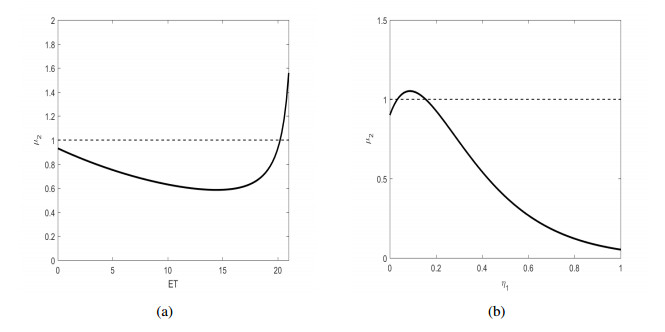
Figures(6)

Yongfeng Li, Song Huang, Zhongyi Xiang. A state-dependent impulsive system with ratio-dependent action threshold for investigating SIR model[J]. AIMS Mathematics, 2024, 9(2): 4781-4804. doi: 10.3934/math.2024231







 DownLoad:
DownLoad: