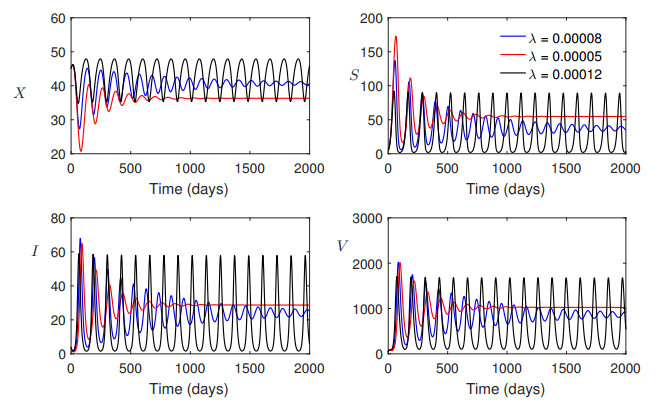
Mathematical modeling and analysis of a crop-pest interacting system helps us to understand the dynamical properties of the system such as stability, bifurcations and chaos. In this article, a predator-prey type mathematical model for pest control using bio-pesticides has been analysed to study the global stability property of the interior equilibrium point. Moreover, the occurrence and orbital stability of Hopf bifurcating limit cycle solutions have been studied using ref30's conditions. Analytical and numerical results show that the interior equilibrium of the pest control model is globally asymptotically stable. Also, Hopf bifurcating occurs when the bifurcation parameter crosses the critical value, and the bifurcating periodic solution is found to be stable.
Citation: Aeshah A. Raezah, Jahangir Chowdhury, Fahad Al Basir. Global stability of the interior equilibrium and the stability of Hopf bifurcating limit cycle in a model for crop pest control[J]. AIMS Mathematics, 2024, 9(9): 24229-24246. doi: 10.3934/math.20241179
Mathematical modeling and analysis of a crop-pest interacting system helps us to understand the dynamical properties of the system such as stability, bifurcations and chaos. In this article, a predator-prey type mathematical model for pest control using bio-pesticides has been analysed to study the global stability property of the interior equilibrium point. Moreover, the occurrence and orbital stability of Hopf bifurcating limit cycle solutions have been studied using ref30's conditions. Analytical and numerical results show that the interior equilibrium of the pest control model is globally asymptotically stable. Also, Hopf bifurcating occurs when the bifurcation parameter crosses the critical value, and the bifurcating periodic solution is found to be stable.

| [1] | S. Gupta, A. K. Dikshit, Biopesticides: An ecofriendly approach for pest control, J. Biopest, (2010), 186. |
| [2] |
W. J. Lewis, J. C. Van Lenteren, S. C. Phatak, J. H. Tumlinson, A total system approach to sustainable pest management, Proc. Natl. Acad. Sci., 94, (1997), 12243–12248. https://doi.org/10.1073/pnas.94.23.1224 doi: 10.1073/pnas.94.23.1224

|
| [3] | M. L. Flint, R. Van den Bosch, Introduction to integrated pest management, Springer Science & Business Media, (2012). |
| [4] | E. Beltrami, Mathematics for dynamic modeling, Academic press, (2014). |
| [5] | R. M. May, Stability and complexity in model ecosystems, Princeton university press, (2019). |
| [6] |
L. F. Cavalieri, H. Koçak, Chaos in biological control systems, J. Theoret. Biol., 169 (1994), 179–187. https://doi.org/10.1006/jtbi.1994.1139 doi: 10.1006/jtbi.1994.1139
|
| [7] |
W. L. Keith, R. H. Rand, Dynamics of a system exhibiting the global bifurcation of a limit cycle at infinity, Int. J. Non-Lin. Mech., 20 (1985), 325–338. https://doi.org/10.1016/0020-7462(85)90040-X doi: 10.1016/0020-7462(85)90040-X
|
| [8] | S. Sastry, Nonlinear systems: Analysis, stability, and control, Springer Science, Business Media, 10 (2013). |
| [9] | R. Seydel, Practical bifurcation and stability analysis, Springer Science & Business Media, (2009). |
| [10] |
Z. He, X. Lai, Bifurcation and chaotic behavior of a discrete-time predator–prey system, Nonlinear Anal. Real. World Appl., 12 (2019), 403–417. https://doi.org/10.1016/j.nonrwa.2010.06.026 doi: 10.1016/j.nonrwa.2010.06.026
|
| [11] | S. H. Strogatz, Nonlinear dynamics and chaos with student solutions manual: With applications to physics, biology, chemistry, and engineering, CRC press, (2018). |
| [12] |
V. Kumar, J. Dhar, H. S. Bhatti, Stability and Hopf bifurcation dynamics of a food chain system: plant–pest–natural enemy with dual gestation delay as a biological control strategy, Model. Earth Syst. Environ., 4 (2018), 881–889. https://doi.org/10.1007/s40808-018-0417-1 doi: 10.1007/s40808-018-0417-1
|
| [13] |
F. A. Basir, A multi-delay model for pest control with awareness induced interventions—Hopf bifurcation and optimal control analysis, Int. J. Biomath., 13 (2020), 2050047. https://doi.org/10.1142/S1793524520500473 doi: 10.1142/S1793524520500473
|
| [14] |
T. Abraha, F. Al Basir, L. L. Obsu, D. F. M. Torres, Farming awareness based optimum interventions for crop pest control, Math. Biosci. Eng., 18 (2021), 5364–5391. https://doi.org/10.3934/mbe.2021272 doi: 10.3934/mbe.2021272
|
| [15] | W. Costello, H. Taylor, Mathematical models of the sterile male technique of insect control, in: Mathematical Analysis of Decision Problems in Ecology, Springer, Berlin, Heidelberg, (1975), 318–359. https://doi.org/10.1007/978-3-642-80924-8_12 |
| [16] |
T. L. Vincent, Pest management programs via optimal control theory, Biometrics, 31 (1975), 1–10. https://doi.org/10.2307/2529704 doi: 10.2307/2529704
|
| [17] |
Y. Liu, Y. Yang, B. Wang, Entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae play roles of maize (Zea mays) growth promoter, Sci. Rep., 12 (2022), 15706. https://doi.org/10.1038/s41598-022-19899-7 doi: 10.1038/s41598-022-19899-7
|
| [18] |
F. A. Basir, S. Samanta, P. K. Tiwari, Bistability, generalized and zero-hopf bifurcations in a pest control model with farming awareness, J. Biol. Syst., 31 (2023), 115–140. https://doi.org/10.1142/S0218339023500079 doi: 10.1142/S0218339023500079
|
| [19] |
G. Seo, G. S. Wolkowicz, Pest control by generalist parasitoids: A bifurcation theory approach. Discrete Cont. Dyn. S., 31 (2020), 3157–3187. https://doi.org/10.3934/dcdss.2020163 doi: 10.3934/dcdss.2020163
|
| [20] | D. K. Bhattacharya, S. Karan, On bionomic model of integrated pest management of a single pest population, J. Differ. Equat. Dyn. Syst., 12 (2004), 301–330. |
| [21] |
S. Ghosh, D. K. Bhattacharyya, Optimization in microbial pest control: An integrated approach, Appl. Math. Model., 34 (2010), 1382–1395. https://doi.org/10.1016/j.apm.2009.08.026 doi: 10.1016/j.apm.2009.08.026
|
| [22] | F. A. Basir, A. Banerjee, S. Ray, Role of farming awareness in crop pest management—a mathematical model, J. Theoret. Biol., 461 (2019), 59–67. |
| [23] | E. Kurstak, Microbial and Viral Pesticide, Marcel and Dekker, Inc., New York, Bessel, (1982). |
| [24] |
S. Bhattacharyya, D. K. Bhattacharyya, An improved integrated pest management model under 2-control parameters (sterile male and pesticide), Math. Biosci., 209, (2007), 256–281. https://doi.org/10.1016/j.mbs.2006.08.003 doi: 10.1016/j.mbs.2006.08.003
|
| [25] | J. Chowdhury, F. Al Basir, J. Pal, P. K. Roy, Pest control for Jatropha curcas plant through viral disease: a mathematical approach, Nonlinear Stud., 23 (2016), 517–532. |
| [26] |
T. Abraha, F. A. Basir, L. L. Obsu, D. F. M. Torres, Pest control using farming awareness: Impact of time delays and optimal use of biopesticides, Chaos Soliton. Fract., 146 (2021), 110869. https://doi.org/10.1016/j.chaos.2021.110869 doi: 10.1016/j.chaos.2021.110869
|
| [27] |
S. Ghosh, S. Bhattacharyya, D.K. Bhattacharyya, The Role of Viral infection in Pest Control: A Mathematical Study, Bull. Math. Biol., 69 (2007), 2649–2691. https://doi.org/10.1007/s11538-007-9235-8 doi: 10.1007/s11538-007-9235-8
|
| [28] |
J. Chowdhury, F. A. Basir, Y. Takeuchi, M. Ghosh, P. K. Roy, A mathematical model for pest management in Jatropha curcas with integrated pesticides–an optimal control approach, Ecol. Complex., 37 (2019), 24–31. https://doi.org/10.1016/j.ecocom.2018.12.004 doi: 10.1016/j.ecocom.2018.12.004
|
| [29] |
A. B. Poore, On the theory and application of the Hopf-Friedrichs bifurcation theory, Arch. Rat. Mech. Anal., 60 (1976), 371–393. https://doi.org/10.1007/BF00248886 doi: 10.1007/BF00248886
|



Figures(4) / Tables(1)

Aeshah A. Raezah, Jahangir Chowdhury, Fahad Al Basir. Global stability of the interior equilibrium and the stability of Hopf bifurcating limit cycle in a model for crop pest control[J]. AIMS Mathematics, 2024, 9(9): 24229-24246. doi: 10.3934/math.20241179







 DownLoad:
DownLoad: