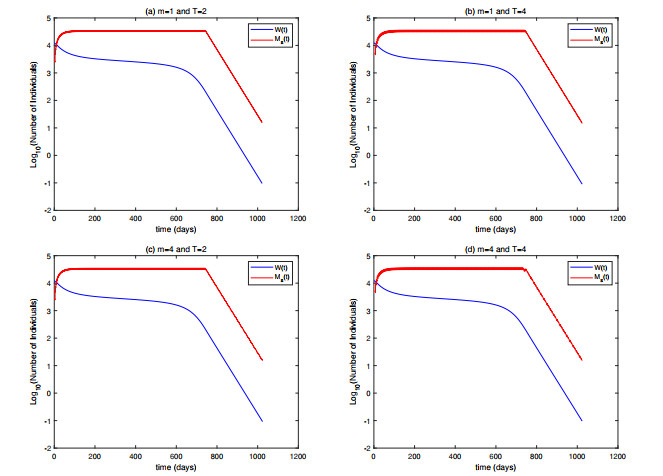
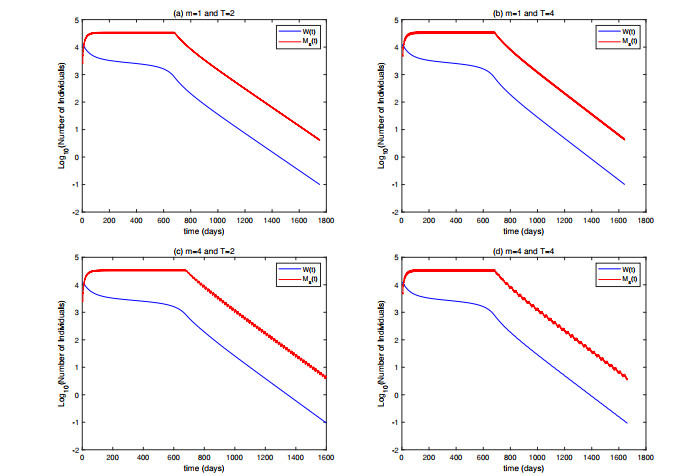
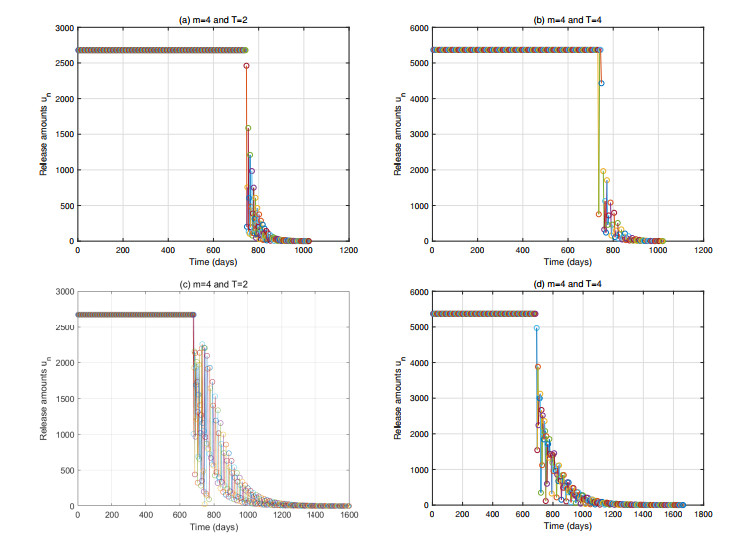
In this paper, we proposed a model for suppressing mosquito populations, evaluating various release strategies for sterilized mosquitoes, including constant continuous release, open-loop control, closed-loop control, and composite control strategies. We established release amount thresholds for each strategy to ensure the ultimate extinction of the wild population. Through numerical simulations, we validated our theoretical results and assessed the efficacy of each strategy. The results indicated that the closed-loop control mechanism significantly enhanced population suppression effectiveness. Higher intensity control notably shortened the control duration, and strategies that incorporated shorter release periods and more frequent population assessments markedly reduced the cumulative release quantity. Furthermore, we found that, compared to closed-loop control, the composite control strategy reduced single release amounts but did not show advantages in other aspects at low control intensities. However, it significantly decreased the cumulative release quantity at high control intensities.
Citation: Shouzong Liu, Yang Xu, Mingzhan Huang. Comparative analysis of sterile mosquito release strategies based on a population suppression model[J]. AIMS Mathematics, 2024, 9(9): 23344-23367. doi: 10.3934/math.20241135
In this paper, we proposed a model for suppressing mosquito populations, evaluating various release strategies for sterilized mosquitoes, including constant continuous release, open-loop control, closed-loop control, and composite control strategies. We established release amount thresholds for each strategy to ensure the ultimate extinction of the wild population. Through numerical simulations, we validated our theoretical results and assessed the efficacy of each strategy. The results indicated that the closed-loop control mechanism significantly enhanced population suppression effectiveness. Higher intensity control notably shortened the control duration, and strategies that incorporated shorter release periods and more frequent population assessments markedly reduced the cumulative release quantity. Furthermore, we found that, compared to closed-loop control, the composite control strategy reduced single release amounts but did not show advantages in other aspects at low control intensities. However, it significantly decreased the cumulative release quantity at high control intensities.

| [1] |
M. Strugarek, H. Bossin, Y. Dumont, On the use of the sterile insect release technique to reduce or eliminate mosquito populations, Appl. Math. Model., 68 (2019), 443–470. http://dx.doi.org/10.1016/j.apm.2018.11.026 doi: 10.1016/j.apm.2018.11.026

|
| [2] |
P. Bliman, D. Cardona-Salgado, Y. Dumont, O. Vasilieva, Implementation of control strategies for sterile insect techniques, Math. Biosci., 314 (2019), 43–60. http://dx.doi.org/10.1016/j.mbs.2019.06.002 doi: 10.1016/j.mbs.2019.06.002
|
| [3] |
R. Lees, J. Gilles, J. Hendrichs, M. Vreysen, K. Bourtzis, Back to the future: the sterile insect technique against mosquito disease vectors, Curr. Opin. Insect Sci., 10 (2015), 156–162. http://dx.doi.org/10.1016/j.cois.2015.05.011 doi: 10.1016/j.cois.2015.05.011
|
| [4] |
L. Cai, S. Ai, J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. http://dx.doi.org/10.1137/13094102X doi: 10.1137/13094102X
|
| [5] |
J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dynam., 11 (2017), 316–333. http://dx.doi.org/10.1080/17513758.2016.1216613 doi: 10.1080/17513758.2016.1216613
|
| [6] |
J. Li, L. Cai, Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dynam., 11 (2017), 79–101. http://dx.doi.org/10.1080/17513758.2016.1159740 doi: 10.1080/17513758.2016.1159740
|
| [7] |
Y. Hui, J. Yu, Global asymptotic stability in a non-autonomous delay mosquito population suppression model, Appl. Math. Lett., 124 (2022), 107599. http://dx.doi.org/10.1016/j.aml.2021.107599 doi: 10.1016/j.aml.2021.107599
|
| [8] |
M. Huang, S. Liu, X. Song, Study of a delayed mosquito population suppression model with stage and sex structure, J. Appl. Math. Comput., 69 (2023), 89–111. http://dx.doi.org/10.1007/s12190-022-01735-w doi: 10.1007/s12190-022-01735-w
|
| [9] |
J. Yu, J. Li, Discrete-time models for interactive wild and sterile mosquitoes with general time steps, Math. Biosci., 346 (2022), 108797. http://dx.doi.org/10.1016/j.mbs.2022.108797 doi: 10.1016/j.mbs.2022.108797
|
| [10] |
Y. Li, J. Li, Stage-structured discrete-time models for interacting wild and sterile mosquitoes with beverton-holt survivability, Math. Biosci. Eng., 16 (2019), 572–602. http://dx.doi.org/10.3934/mbe.2019028 doi: 10.3934/mbe.2019028
|
| [11] |
M. Huang, S. Liu, X. Song, Modeling of periodic compensation policy for sterile mosquitoes incorporating sexual lifespan, Math. Method. Appl. Sci., 46 (2023), 5725–5741. http://dx.doi.org/10.1002/mma.8864 doi: 10.1002/mma.8864
|
| [12] |
M. Huang, S. Liu, X. Song, Study of the sterile insect release technique for a two-sex mosquito population model, Math. Biosci. Eng., 18 (2021), 1314–1339. http://dx.doi.org/10.3934/mbe.2021069 doi: 10.3934/mbe.2021069
|
| [13] |
B. Zheng, J. Yu, J. Li, Existence and stability of periodic solutions in a mosquito population suppression model with time delay, J. Differ. Equations, 315 (2022), 159–178. http://dx.doi.org/10.1016/j.jde.2022.01.036 doi: 10.1016/j.jde.2022.01.036
|
| [14] |
B. Zheng, J. Yu, At most two periodic solutions for a switching mosquito population suppression model, J. Dyn. Diff. Equat., 35 (2023), 2997–3009. http://dx.doi.org/10.1007/s10884-021-10125-y doi: 10.1007/s10884-021-10125-y
|
| [15] |
M. Huang, X. Song, J. Li, Modelling and analysis of impulsive release of sterile mosquitoes, J. Biol. Dynam., 11 (2017), 147–171. http://dx.doi.org/10.1080/17513758.2016.1254286 doi: 10.1080/17513758.2016.1254286
|
| [16] |
G. Jiang, Q. Lu, L. Qian, Complex dynamics of a Holling type Ⅱ prey-predator system with state feedback control, Chaos Soliton. Fract., 31 (2007), 448–461. http://dx.doi.org/10.1016/j.chaos.2005.09.077 doi: 10.1016/j.chaos.2005.09.077
|
| [17] |
Q. Zhang, S. Tang, X. Zou, Rich dynamics of a predator-prey system with state-dependent impulsive controls switching between two means, J. Differ. Equations, 364 (2023), 336–377. http://dx.doi.org/10.1016/j.jde.2023.03.030 doi: 10.1016/j.jde.2023.03.030
|
| [18] |
J. Yang, G. Tang, S. Tang, Holling-Tanner predator-prey model with state-dependent feedback control, Discrete Dyn. Nat. Soc., 2018 (2018), 3467405. http://dx.doi.org/10.1155/2018/3467405 doi: 10.1155/2018/3467405
|
| [19] |
H. Xu, T. Zhang, H. Cheng, Nonlinear control ecological model with complex discrete map, Commun. Nonlinear Sci., 118 (2023), 107019. http://dx.doi.org/10.1016/j.cnsns.2022.107019 doi: 10.1016/j.cnsns.2022.107019
|
| [20] |
Y. Tian, Y. Gao, K. Sun, Global dynamics analysis of instantaneous harvest fishery model guided by weighted escapement strategy, Chaos Soliton. Fract., 164 (2022), 112597. http://dx.doi.org/10.1016/j.chaos.2022.112597 doi: 10.1016/j.chaos.2022.112597
|
| [21] |
Y. Tian, Y. Gao, K. Sun, Qualitative analysis of exponential power rate fishery model and complex dynamics guided by a discontinuous weighted fishing strategy, Commun. Nonlinear Sci., 118 (2023), 107011. http://dx.doi.org/10.1016/j.cnsns.2022.107011 doi: 10.1016/j.cnsns.2022.107011
|
| [22] |
J. Jia, Z. Zhao, J. Yang, A. Zeb, Parameter estimation and global sensitivity analysis of a bacterial-plasmid model with impulsive drug treatment, Chaos Soliton. Fract., 183 (2024), 114901. http://dx.doi.org/10.1016/j.chaos.2024.114901 doi: 10.1016/j.chaos.2024.114901
|
| [23] |
M. Huang, J. Li, X. Song, H. Guo, Modeling impulsive injections of insulin: towards artificial pancreas, SIAM J. Appl. Math., 72 (2012), 1524–1548. http://dx.doi.org/10.1137/110860306 doi: 10.1137/110860306
|
| [24] |
L. O'Connor, C. Plichart, A. Sang, C. Brelsfoard, H. Bossin, S. Dobson, Open release of male mosquitoes infected with a Wolbachia biopesticide: field performance and infection containment, PLOS Negl. Trop. Dis., 6 (2012), 1797. http://dx.doi.org/10.1371/journal.pntd.0001797 doi: 10.1371/journal.pntd.0001797
|

Figures(8) / Tables(4)

Shouzong Liu, Yang Xu, Mingzhan Huang. Comparative analysis of sterile mosquito release strategies based on a population suppression model[J]. AIMS Mathematics, 2024, 9(9): 23344-23367. doi: 10.3934/math.20241135







 DownLoad:
DownLoad: