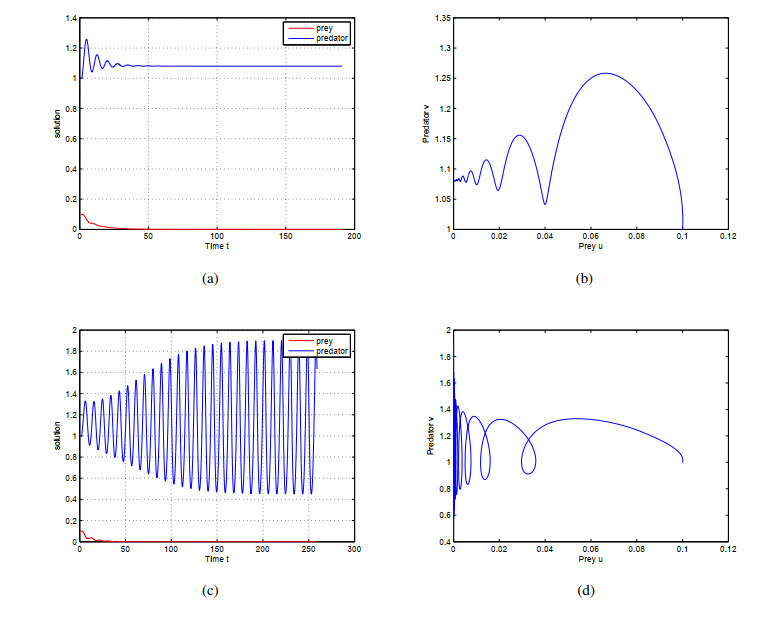
In this paper, a delayed predator-prey system with additional food and asymmetric functional response is investigated. We discuss the local stability of equilibria and the existence of local Hopf bifurcation under the influence of the time delay. By using the normal form theory and center manifold theorem, the explicit formulas which determine the properties of bifurcating periodic solutions are obtained. Further, we prove that global periodic solutions exist after the second critical value of delay via Wu's theory. Finally, the correctness of the previous theoretical analysis is demonstrated by some numerical cases.
Citation: Luoyi Wu, Hang Zheng. Hopf bifurcation in a delayed predator-prey system with asymmetric functional response and additional food[J]. AIMS Mathematics, 2021, 6(11): 12225-12244. doi: 10.3934/math.2021708
In this paper, a delayed predator-prey system with additional food and asymmetric functional response is investigated. We discuss the local stability of equilibria and the existence of local Hopf bifurcation under the influence of the time delay. By using the normal form theory and center manifold theorem, the explicit formulas which determine the properties of bifurcating periodic solutions are obtained. Further, we prove that global periodic solutions exist after the second critical value of delay via Wu's theory. Finally, the correctness of the previous theoretical analysis is demonstrated by some numerical cases.

| [1] |
S. Tang, L. Chen, Global attractivity in a "food-limited" population model with impulsive effects, J. Math. Anal. Appl., 292 (2004), 211–221. doi: 10.1016/j.jmaa.2003.11.061

|
| [2] |
G. Saunders, B. Cooke, K. McColl, R. Shine, T. Peacock, Modern approaches for the biological control of vertebrate pests: an Australian perspective, Biol. Control, 52 (2010), 288–295. doi: 10.1016/j.biocontrol.2009.06.014
|
| [3] | B. Ghosh, F. Grognard, L. Mailleret, Natural enemies deployment in patchy environments for augmentative biological control, Appl. Math. Comput., 266 (2015), 982–999. |
| [4] |
A. Tena, A. Pekas, D. Cano, F. Wackers, A. Urbaneja, Sugar provisioning maximizes the biocontrol service of parasitoids, J. Appl. Ecol., 52 (2015), 795–804. doi: 10.1111/1365-2664.12426
|
| [5] |
P. Srinivasu, B. Prasad, M. Venkatesulu, Biological control through provision of additional food to predators: a theoretical study, Theor. Popul. Biol., 72 (2007), 111–120. doi: 10.1016/j.tpb.2007.03.011
|
| [6] |
B. Sahoo, S. Poria, The chaos and control of a food chain model supplying additional food to top-predator, Chaos Soliton. Fract., 58 (2014), 52–64. doi: 10.1016/j.chaos.2013.11.008
|
| [7] |
T. Kar, B. Ghosh, Sustainability and optimal control of an exploited prey predator system through provision of alternative food to predator, Biosystems, 109 (2012), 220–232. doi: 10.1016/j.biosystems.2012.02.003
|
| [8] |
P. Srinivasu, B. Prasad, Role of quantity of additional food to predators as a control in predator prey systems with relevance to pest management and biological conservation, Bull Math. Biol., 73 (2011), 2249–2276. doi: 10.1007/s11538-010-9601-9
|
| [9] | J. Song, M. Hu, Y. Bai, Y. Xia, Dynamic analysis of a non-autonomous ratio-dependent predator-prey model with additional food, J. Appl. Anal. Comput., 8 (2018), 1893–1909. |
| [10] | B. Sahoo, S. Poria, Effects of additional food on an ecoepidemic model with time delay on infection, Appl. Math. Comput., 245 (2014), 17–35. |
| [11] | B. Sahoo, Role of additional food in eco-epidemiological system with disease in the prey, Appl. Math. Comput., 259 (2015), 61–79. |
| [12] |
A. Basheer, E. Quansah, R. Parshad, The effect of additional food in Holling Tanner type models, Int. J. Dyn. Control, 7 (2019), 1195–1212. doi: 10.1007/s40435-019-00580-3
|
| [13] |
Y. Xia, Global analysis of an impulsive delayed Lotka-Volterra competition system, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 1597–1616. doi: 10.1016/j.cnsns.2010.07.014
|
| [14] | X. Meng, L. Chen, Permanence and global stability in an impulsive lotka-volterra n-species competitive system with both discrete delays and continuous delays, Int. J. Biomath., 1 (2012), 179–196. |
| [15] |
X. Meng, L. Chen, Almost periodic solution of non-autonomous Lotka-Volterra predator-prey dispersal system with delays, J. Theoret. Biol., 243 (2006), 562–574. doi: 10.1016/j.jtbi.2006.07.010
|
| [16] |
Z. Wei, Y. Xia, T. Zhang, Stability and bifurcation analysis of a amensalism model with weak Allee effect, Qual. Theory Dyn. Syst., 19 (2020), 23. doi: 10.1007/s12346-020-00341-0
|
| [17] |
K. Kou, Y. Lou, Y. Xia, Zeros of a Class of Transcendental Equation with Application to Bifurcation of DDE, Int. J. Bifurcat. Chaos, 26 (2016), 1650062. doi: 10.1142/S0218127416500620
|
| [18] |
R. Xu, Z. Ma, Stability and Hopf bifurcation in a ratio-dependent predator-prey system with stage structure, Chaos Soliton. Fract., 38 (2008), 669–684. doi: 10.1016/j.chaos.2007.01.019
|
| [19] |
A. Maiti, B. Dubey, A. Chakraborty, Global analysis of a delayed stage structure prey-predator model with Crowley-Martin type functional response, Math. Comput. Simulat., 162 (2019), 58–84. doi: 10.1016/j.matcom.2019.01.009
|
| [20] |
Y. Chen, F. Zhang, Dynamics of a delayed predator-prey model with predator migration, Appl. Math. Model., 37 (2013), 1400–1412. doi: 10.1016/j.apm.2012.04.012
|
| [21] | T. Kar, U. Das, Bifurcation analysis of a delayed predator-prey model with Holling type III functional response and predator harvesting, J. Nonlinear Dyn., 2014 (2014), 1–10. |
| [22] | J. Jiao, Y. Song, Stability and bifurcation analysis of a delayed Leslie-Gower predator-prey system with nonmonotonic functional response, Abstr. Appl. Anal., 2013 (2013), 152459. |
| [23] |
Y. Song, Y. Tao, H. Shu, Dynamics of a ratio-dependent stage-structured predator-prey model with delay, Math. Methods Appl. Sci., 40 (2017), 6451–6467. doi: 10.1002/mma.4467
|
| [24] | X. Jiang, X. Chen, T. Huang, H. Yan, Bifurcation and Control for a Predator-Prey System With Two Delays, IEEE T. Circuits-II, 68 (2021), 376–380. |
| [25] | X. Jiang, X. Chen, M. Chi, J. Chen, On Hopf bifurcation and control for a delay systems, Appl. Math. Comput., 370 (2019), 124906. |
| [26] |
B. Barman, B. Ghosh, Explicit impacts of harvesting in delayed predator-prey models, Chaos Soliton. Fract., 122 (2019), 213–228. doi: 10.1016/j.chaos.2019.03.002
|
| [27] |
L. Erbe, K. Geba, W. Krawcewicz, J. Wu, $S^1$-degree and global Hopf bifurcations, J. Differ. Equations, 98 (1992), 277–298. doi: 10.1016/0022-0396(92)90094-4
|
| [28] |
J. Wu, Symmetric functional differential equations and neural networks with memory, Trans. Amer. Math. Soc., 350 (1998), 4799–4838. doi: 10.1090/S0002-9947-98-02083-2
|
| [29] |
Y. Song, J. Wei, Local Hopf bifurcation and global periodic solutions in a delayed predator-prey system, J. Math. Anal. Appl., 301 (2005), 1–21. doi: 10.1016/j.jmaa.2004.06.056
|
| [30] |
S. Yuan, F. Zhang, Stability and global Hopf bifurcation in a delayed predator-prey system, Nonlinear Anal. Real World Appl., 11 (2010), 959–977. doi: 10.1016/j.nonrwa.2009.01.038
|
| [31] |
S. Yuan, Y. Song, Stability and Hopf bifurcations in a delayed Leslie-Gower predator-prey system, J. Math. Anal. Appl., 355 (2009), 82–100. doi: 10.1016/j.jmaa.2009.01.052
|
| [32] | P. Hao, X. Wang, J. Wei, Global Hopf bifurcation of a population model with stage structure and strong Allee effect, Discrete Contin. Dyn. Syst. Ser. S, 10 (2017), 973–993. |
| [33] | X. Yan, W. Li, Hopf bifurcation and global periodic solutions in a delayed predator-prey system, Appl. Math. Comput., 177 (2006), 427–445. |
| [34] |
X. Liu, T. Zhang, X. Meng, T. Zhang, Turing-Hopf bifurcations in a predator-prey model with herd behavior, quadratic mortality and prey-taxis, Phys. A, 496 (2018), 446–460. doi: 10.1016/j.physa.2018.01.006
|
| [35] | B. Hassard, N. Kazarinoff, Y. Wan, Theory and Applications of Hopf Bifurcation, Cambridge: Cambridge Univ. Press, 1981. |



Figures(4)

Luoyi Wu, Hang Zheng. Hopf bifurcation in a delayed predator-prey system with asymmetric functional response and additional food[J]. AIMS Mathematics, 2021, 6(11): 12225-12244. doi: 10.3934/math.2021708







 DownLoad:
DownLoad: