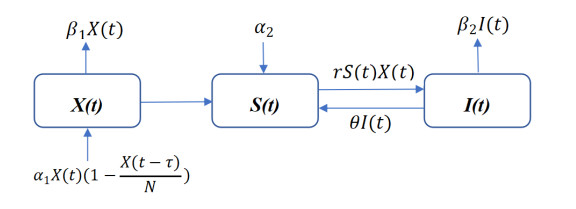
In this paper, we establish an infectious disease model of Tropidothorax elegans to study the impact of them on plants. Our model involves the time delay for Tropidothorax elegans to hatch eggs, which is influenced by temperature. Second, we theoretically analyze the existence and the stability of the equilibrium and the normal form near the Hopf bifurcating critical point. Next, we choose three groups of parameters for numerical simulations to verify theoretical analysis of our model. Then, based on numerical simulations, we give bioanalysis which are consistent with the patterns of Tropidothorax elegans pests, such as dying off in large numbers of adults during the winter and one or two generations a year.
Citation: Tingru Yang, Yuting Ding. Dynamic analysis of a delayed differential equation for Tropidothorax elegans pests[J]. Electronic Research Archive, 2023, 31(11): 6947-6963. doi: 10.3934/era.2023352
In this paper, we establish an infectious disease model of Tropidothorax elegans to study the impact of them on plants. Our model involves the time delay for Tropidothorax elegans to hatch eggs, which is influenced by temperature. Second, we theoretically analyze the existence and the stability of the equilibrium and the normal form near the Hopf bifurcating critical point. Next, we choose three groups of parameters for numerical simulations to verify theoretical analysis of our model. Then, based on numerical simulations, we give bioanalysis which are consistent with the patterns of Tropidothorax elegans pests, such as dying off in large numbers of adults during the winter and one or two generations a year.

| [1] |
W. Yang, Z. Min, M. X. Yang, J. Yan, Exploration of the implementation of carbon neutralization in the field of natural resources under the background of sustainable development-an overview, Int. J. Environ. Res. Public Health, 19 (2022), 14109. https://doi.org/10.3390/ijerph192114109 doi: 10.3390/ijerph192114109

|
| [2] |
H. Chen, C. C. Tie, Y. Teng, Z. Geng, J. M. Zhu, Research on forest carbon sequestration assessment and forest management plan based on TOPSIS method and grey correlation analysis, Int. J. Environ. Pollut., 70 (2021), 130–147. https://doi.org/10.1504/IJEP.2021.132005 doi: 10.1504/IJEP.2021.132005
|
| [3] |
B. Yang, W. Q. Cao, C. M. Tian, Visual analysis of impact factors of forest pests and diseases, J. Visualization, 22 (2019), 1257–1280. https://doi.org/10.1007/s12650-019-00593-2 doi: 10.1007/s12650-019-00593-2
|
| [4] |
X. H. Yao, Y. P. Xu, B. S. Yuan, L. Y. Han, X. L. Yao, H. H. Han, Study of meteorological assessment techniques of forest diseases and pests in Xiaolong mountain (in Chinese), J. Northwest A, F Univ. (Nat. Sci. Ed.), 10 (2012), 64–68. https://doi.org/10.13207/j.cnki.jnwafu.2012.10.018 doi: 10.13207/j.cnki.jnwafu.2012.10.018
|
| [5] |
D. Kiewra, M. Kryza, M. Szymanowski, Influence of selected meteorological variables on the questing activity of Ixodes ricinus ticks in Lower Silesia, SW Poland, J. Vector Ecol., 39 (2014), 138–145. https://doi.org/10.1111/j.1948-7134.2014.12080.x doi: 10.1111/j.1948-7134.2014.12080.x
|
| [6] |
W. Wang, W. T. Peng, X. Y. Liu, G. He, Y. L. Cai, Spatiotemporal dynamics and factors driving the distributions of pine wilt disease-damaged forests in China, Forests, 13 (2022), 261. https://doi.org/10.3390/f13020261 doi: 10.3390/f13020261
|
| [7] | R. Castro, S. Lanucara, V. Piccione, G. Pioggia, G. Modica, M. A. Ragusa, MEDALUS model evolutions and prospects case study Sicily, in International Conference on Computational Science and Its Applications, Springer Nature Switzerland, (2023), 310–326. https://doi.org/10.1007/978-3-031-37114-1_21 |
| [8] |
X. M. Chen, D. M. Luo, Dynamical analysis of an almost periodic multispecies mutualism system with impulsive effects and time delays, Filomat, 37 (2023), 551–565. https://doi.org/10.2298/FIL2302551C doi: 10.2298/FIL2302551C
|
| [9] |
Z. Khan, J. Khan, S. Saifullah, A. Ali, Dynamics of hidden attractors in fourdimensional dynamical systems with power law, J. Funct. Spaces, 2022 (2022), 3675076. https://doi.org/10.1155/2022/3675076 doi: 10.1155/2022/3675076
|
| [10] |
H. A. Wood, R. R. Granados, Genetically engineered baculoviruses as agents for pest control, Annu. Rev. Microbiol., 45 (1991), 69–87. https://doi.org/10.1146/annurev.mi.45.100191.000441 doi: 10.1146/annurev.mi.45.100191.000441
|
| [11] |
V. Mondello, A. Songy, E. Battison, C. Pinto, C. Coppin, P. Trozel-Aziz, et al., Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents, Plant Dis., 102 (2018), 1189–1217. https://doi.org/10.1094/PDIS-08-17-1181-FE doi: 10.1094/PDIS-08-17-1181-FE
|
| [12] |
T. Shimales, E. Mendesil, B. Zewdie, B. Ayalew, K. Hylander, A. J. M. Tack, Management intensity affects insect pests and natural pest control on Arabica coffee in its native range, J. Appl. Ecol., 60 (2023), 911–922. https://doi.org/10.1111/1365-2664.14410 doi: 10.1111/1365-2664.14410
|
| [13] |
W. X. Yao, S. Guo, J. Wang, C. L. Chen, F. H. Yu, X. Li, et al., Droplet deposition and pest control efficacy on pine trees from aerial application, Pest. Manage. Sci., 78 (2022), 3324–3336. https://doi.org/10.1002/ps.6959 doi: 10.1002/ps.6959
|
| [14] |
Z. Arslangundogdu, E. Hizal, S. Acer, First record of oxycarenus lavaterae (Fabricius, 1787) (Heteroptera, Lygaeidae) in Turkey, Appl. Ecol. Environ. Res., 16 (2018), 1305–1311. https://doi.org/10.15666/aeer/1602_13051311 doi: 10.15666/aeer/1602_13051311
|
| [15] | C. Q. Yang, Y. Fan, C. G. Du, A preliminary study on the Tropidothorax elegans (in Chinese), Insect Knowl., 25 (1988), 36–38. |
| [16] |
W. S. Avusuglo, R. Mosleh, T. Ramaj, A. Li, S. S. Sharbayta, A. A. Fall, et al., Workplace absenteeism due to COVID-19 and influenza across Canada: a mathematical model, J. Theor. Biol., 572 (2023), 111559. https://doi.org/10.1016/j.jtbi.2023.111559 doi: 10.1016/j.jtbi.2023.111559
|
| [17] |
L. L. Chang, W. Gong, Z. Jin, G. Q. Sun, Sparse optimal control of pattern formations for an SIR reaction-diffusion epidemic model, SIAM J. Appl. Math., 82 (2022), 1764–1790. https://doi.org/10.1137/22M1472127 doi: 10.1137/22M1472127
|
| [18] |
Y. Tatsukawa, M. R. Arefin, S. Utsumi, K. Kuga, J. Tanimoto, Stochasticity of disease spreading derived from the microscopic simulation approach for various physical contact networks, Appl. Math. Comput., 431 (2022), 127328. https://doi.org/10.1016/j.amc.2022.127328 doi: 10.1016/j.amc.2022.127328
|
| [19] |
G. Q. Sun, X. Ma, Z. Z. Zhang, Q. H. Liu, B. L. Li, What is the role of aerosol transmission in SARS-Cov-2 Omicron spread in Shanghai, BMC Infect. Dis., 22 (2022), 880. https://doi.org/10.1186/s12879-022-07876-4 doi: 10.1186/s12879-022-07876-4
|
| [20] |
X. Ma, G. Q. Sun, Z. H. Wang, Y. M. Chu, Z. Jin, B. L. Li, Transmission dynamics of brucellosis in Jilin province, China: Effects of different control measures, Commun. Nonlinear Sci., 114 (2022), 106702. https://doi.org/10.1016/j.cnsns.2022.106702 doi: 10.1016/j.cnsns.2022.106702
|
| [21] |
G. Q. Sun, H. T. Zhang, L. L. Chang, Z. Jin, H. Wang, S. Q. Ruan, On the dynamics of a diffusive foot-and-mouth disease model with nonlocal infections, SIAM J. Appl. Math., 82 (2022), 1587–1610. https://doi.org/10.1137/21M1412992 doi: 10.1137/21M1412992
|
| [22] |
T. A. Biala, Y. O. Afolabi, A. Q. M. Khaliq, How efficient is contact tracing in mitigating the spread of COVID-19? A mathematical modeling approach, Appl. Math. Model., 103 (2022), 714–730. https://doi.org/10.1016/j.apm.2021.11.011 doi: 10.1016/j.apm.2021.11.011
|
| [23] |
C. A. Deutsch, J. J. Tewksbury, M. Tigchelaar, D. S. Battisti, S. C. Merrill, R. B. Huey, et al., Increase in crop losses to insect pests in a warming climate, Science, 361 (2018), 916–919. https://doi.org/10.1126/science.aat3466 doi: 10.1126/science.aat3466
|
| [24] |
H. Puebla, P. K. Roy, A. Velasco-Perez, M. M. Gonzalez-Brambila, Biological pest control using a model-based robust feedback, IET Syst. Biol, 12 (2018), 233–240. https://doi.org/10.1049/iet-syb.2018.5010 doi: 10.1049/iet-syb.2018.5010
|
| [25] |
L. James, F. D. Hugh, Interspecific hybridization and genetic divergence in Milkweed Bugs (Oncopeltus: Hemiptera: Lygaeidae), Evolution, 37 (1983), 583–591. https://doi.org/10.1111/j.1558-5646.1983.tb05575.x doi: 10.1111/j.1558-5646.1983.tb05575.x
|
| [26] |
A. Duarte, N. Borralho, P. Cabral, M. Caetano, Recent advances in forest insect pests and diseases monitoring using UAV-Based data: A systematic review, Forests, 13 (2022), 911. https://doi.org/10.3390/f13060911 doi: 10.3390/f13060911
|
| [27] |
L. Z. Ji, Z. Wang, X. W. Wang, Forest insect pest management and forest management in China: An overview, Environ. Manage., 48 (2011), 1107–1121. https://doi.org/10.1007/s00267-011-9697-1 doi: 10.1007/s00267-011-9697-1
|
| [28] |
J. N. Liu, Q. Qi, B. Liu, S. J. Gao, Pest control switching models with instantaneous and non-instantaneous impulsive effects, Math. Comput. Simul., 205 (2023), 926–938. https://doi.org/10.1016/j.matcom.2022.10.027 doi: 10.1016/j.matcom.2022.10.027
|





Figures(6) / Tables(1)
Tingru Yang, Yuting Ding. Dynamic analysis of a delayed differential equation for Tropidothorax elegans pests[J]. Electronic Research Archive, 2023, 31(11): 6947-6963. doi: 10.3934/era.2023352







 DownLoad:
DownLoad: