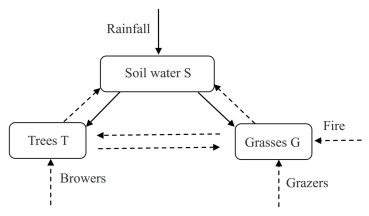
In this work, we consider the dynamic properties of a class of hydrological model with time delay under fire disturbance. The stability of the equilibrium for the model, and the existence of the Hopf bifurcation are analyzed. Moreover, the direction of the Hopf bifurcation, and the stability of these periodic solutions bifurcating are derived based on the normal form and the center manifold theory. Then, the sensitivities of fire intensity and fire frequency to soil water, trees, and grasses are analyzed by the Runge-Kutta method. The result is that, fire frequency has a more significant effect on the hydrological and ecological cycle compared with fire intensity. Finally, we analyze the effect of time delay on the hydrological model through numerical simulations.
Citation: Rina Su. Dynamic analysis for a class of hydrological model with time delay under fire disturbance[J]. Electronic Research Archive, 2022, 30(9): 3290-3319. doi: 10.3934/era.2022167
In this work, we consider the dynamic properties of a class of hydrological model with time delay under fire disturbance. The stability of the equilibrium for the model, and the existence of the Hopf bifurcation are analyzed. Moreover, the direction of the Hopf bifurcation, and the stability of these periodic solutions bifurcating are derived based on the normal form and the center manifold theory. Then, the sensitivities of fire intensity and fire frequency to soil water, trees, and grasses are analyzed by the Runge-Kutta method. The result is that, fire frequency has a more significant effect on the hydrological and ecological cycle compared with fire intensity. Finally, we analyze the effect of time delay on the hydrological model through numerical simulations.

| [1] |
A. Synodinos, B. Tietjen, D. Lohmanna, F. Jeltsch, The impact of inter-annual rainfall variability on African savannas changes with mean rainfall, J. Theor. Biol., 437 (2018), 92–100. https://doi.org/10.1016/j.jtbi.2017.10.019 doi: 10.1016/j.jtbi.2017.10.019

|
| [2] |
R. Su, C. Zhang, Pattern dynamical behaviors of one type of Tree-grass model with cross-diffusion, Int. J. Bifurcation Chaos, 32 (2022), 2250051. https://doi.org/10.1142/S0218127422500511 doi: 10.1142/S0218127422500511
|
| [3] |
M. Sankaran, N. Hanan, R. Scholes, J. Ratnam, D. J. Augustine, B. S. Cade, et al., Determinants of woody cover in African savannas, Nature, 438 (2005), 846–849. https://doi.org/10.1038/nature04070 doi: 10.1038/nature04070
|
| [4] |
P. Amarasekare, Competitive coexistence in spatially structured environments: A synthesis, Ecol. Lett., 6 (2003), 1109–1122. https://doi.org/10.1046/j.1461-0248.2003.00530.x doi: 10.1046/j.1461-0248.2003.00530.x
|
| [5] |
R. Scholes, B. Walker, An African aavanna: Synthesis of the Nylsvley study, J. Appl. Ecol., 31 (1994), 791–792. https://doi.org/10.2307/2404175 doi: 10.2307/2404175
|
| [6] | I. Rodriguez, A. Porporato, Ecohydrology of Water-Controlled Ecosystems: Plant Water Stress, Cambridge University Press, 2005. https://doi.org/10.1017/CBO9780511535727 |
| [7] |
T. Faria, L. Magalhaes, Normal forms for retarded functional differential equations and applications to Bogdanov-Takens singularity, J. Differ. Equations, 122 (1995), 201–224. https://doi.org/10.1006/jdeq.1995.1145 doi: 10.1006/jdeq.1995.1145
|
| [8] |
F. Accatino, C. Michele, R. Vezzoli, D. Donzelli, R. J. Scholes, Tree-grass Co-existence in Savanna: Interactions of rain and fire, J. Theor. Biol., 267 (2010), 235–242. https://doi.org/10.1016/j.jtbi.2010.08.012 doi: 10.1016/j.jtbi.2010.08.012
|
| [9] |
C. Michele, F. Accatino, R. Vezzoli, R. J. Scholes, Savanna domain in the herbivores-fire parameter space exploiting a tree-grass-soil water dynamic model, J. Theor. Biol., 289 (2011), 74–82. https://doi.org/10.1016/j.jtbi.2011.08.014 doi: 10.1016/j.jtbi.2011.08.014
|
| [10] | K. Gopalsamy, Stability and Oscillations in Delay Differential Equations of Population Dynamics, Springer, 1992. https://doi.org/10.1007/978-94-015-7920-9 |
| [11] |
D.Kalyan, R. Santanu, Effect of delay on nutrient cycling in phytoplankton-zooplankton interactions in estuarine system, Ecol. Modell., 215 (2008), 69–76. https://doi.org/10.1016/j.ecolmodel.2008.02.019 doi: 10.1016/j.ecolmodel.2008.02.019
|
| [12] | B. Hassard, N. Kazarinoff, Y. Wan, Theory and applications of Hopf bifurcation, Cambridge University Press, 1981. https://doi.org/10.1002/zamm.19820621221 |
| [13] | A. Pazy, Semigroups of Linear Operators and Applications to Partial Differential Equations, Springer, 1983. https://doi.org/10.1007/978-1-4612-5561-1 |
| [14] |
X. Wu, L. Wang, Zero-Hopf singularity for general delayed differential equations, Nonlinear Dyn., 75 (2014), 141–155. https://doi.org/10.1007/s11071-013-1055-9 doi: 10.1007/s11071-013-1055-9
|
| [15] | J. Hale, S. Chow, Methods of Bifurcation Theory, Springer, 1982. https://doi.org/10.1007/978-1-4613-8159-4 |
| [16] |
T. Min, Y. Cheng, M. Gu, H. You, Parameter estimation of nonlinear dynamic system and sensitivity, Comput. Eng. Appl., 49 (2013), 47–49. 10.3778/j.issn.1002-8331.1110-0488 doi: 10.3778/j.issn.1002-8331.1110-0488
|
| [17] |
S. Higgins, W. Bond, W. Trollope, Fire, resprouting and variability: A recipe for Grass-tree coexistence in Savanna, J. Ecol., 88 (2000), 213–229. https://doi.org/10.1046/j.1365-2745.2000.00435.x doi: 10.1046/j.1365-2745.2000.00435.x
|
| [18] |
A. Synodinos, B. Tietjen, F. Jeltsch, Facilitation in drylands: Modeling a neglected driver of savanna dynamics, Ecol. Modell., 304 (2015), 11–21. https://doi.org/10.1016/j.ecolmodel.2015.02.015 doi: 10.1016/j.ecolmodel.2015.02.015
|
| [19] | B. Luo, Effectis of forest fire disturbance on carbon pools of subtropical forest ecosystem in Guangdong Province, China, Northeast Forestry University, 2020. |











Figures(12) / Tables(2)
Rina Su. Dynamic analysis for a class of hydrological model with time delay under fire disturbance[J]. Electronic Research Archive, 2022, 30(9): 3290-3319. doi: 10.3934/era.2022167







 DownLoad:
DownLoad: