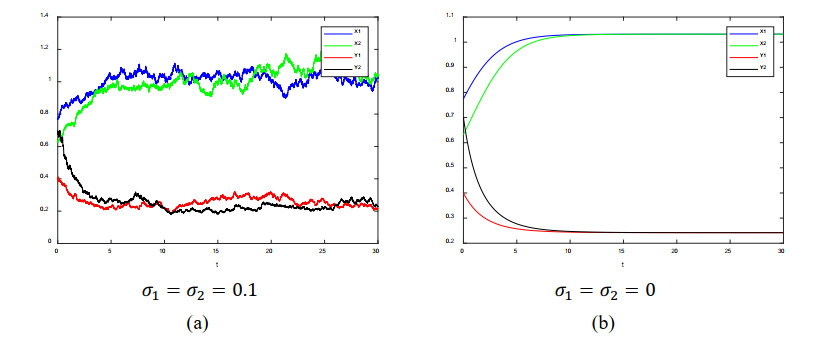
In this paper, a stochastic predator-prey system with mutual interference is studied, which provides guidance on creating appropriate biodegradable environments. By utilizing Mawhin's coincidence degree theorem and constructing a suitable Lyapunov function, a Volterra model with mutual interference is studied. A few sufficient conditions are obtained for existence, extinction and global asymptomatic stability of the positive solution of the model. Then we perform various numerical simulations to verify the stochastic and deterministic systems are global asymptotically stable. It is significant that such a model is firstly proposed with stochastic mutual interference.
Citation: Pinglan Wan. Dynamic behavior of stochastic predator-prey system[J]. Electronic Research Archive, 2023, 31(5): 2925-2939. doi: 10.3934/era.2023147
In this paper, a stochastic predator-prey system with mutual interference is studied, which provides guidance on creating appropriate biodegradable environments. By utilizing Mawhin's coincidence degree theorem and constructing a suitable Lyapunov function, a Volterra model with mutual interference is studied. A few sufficient conditions are obtained for existence, extinction and global asymptomatic stability of the positive solution of the model. Then we perform various numerical simulations to verify the stochastic and deterministic systems are global asymptotically stable. It is significant that such a model is firstly proposed with stochastic mutual interference.

| [1] |
M. Brogi, V. Lagasio, Environmental, social, and governance and company profitability: Are financial intermediaries different?, Corporate Soc. Responsib. Environ. Manage., 26 (2019), 576–587. https://doi.org/10.1002/csr.1704 doi: 10.1002/csr.1704

|
| [2] |
M. A. White, Environmental finance: Value and risk in an age of ecology, Bus. Strategy Environ., 5 (1996), 198–206. https://doi.org/10.1002/(SICI)1099-0836(199609)5:3<198::AID-BSE66>3.0.CO;2-4 doi: 10.1002/(SICI)1099-0836(199609)5:3<198::AID-BSE66>3.0.CO;2-4
|
| [3] |
M. V. Suarez, V. F. Cruz, H. L. Tavera, Biodegradation of polyacrylic and polyester polyurethane coatings by enriched microbial communities, Appl. Microbiol. Biotechnol., 103 (2019), 3225–3236. https://doi.org/10.1007/s00253-019-09660-y doi: 10.1007/s00253-019-09660-y
|
| [4] |
F. Kawai, The biochemistry and molecular biology of xenobiotic polymer degradation by microorganisms, Biosci. Biotechnol. Biochem., 74 (2010), 1743–1759. https://doi.org/10.1271/bbb.100394 doi: 10.1271/bbb.100394
|
| [5] |
E. Nikolaivits, Progressing plastics circularity: A review of mechano-biocatalytic approaches for waste plastic (Re) valorization, Front. Bioeng. Biotechnol., 9 (2021). https://doi.org/10.3389/fbioe.2021.696040 doi: 10.3389/fbioe.2021.696040
|
| [6] |
H. Li, L. Zhang, Z. Teng, Y. Jiang, Dynamic behaviors of holling type ii predator-prey system with mutual interference and impulses, Discrete Dyn. Nat. Soc., 2014 (2014), https://doi.org/10.1155/2014/793761 doi: 10.1155/2014/793761
|
| [7] |
H. Liu, F. Meng, Existence of positive periodic solutions for a predator-prey system of holling type iv function response with mutual interference and impulsive effects, Discrete Dyn. Nat. Soc., 2015 (2015), 1–12. https://doi.org/10.1155/2015/138984 doi: 10.1155/2015/138984
|
| [8] |
K. Wang, Permanence and global asymptotical stability of a predator-prey model with mutual interference, Nonlinear Anal. Real World Appl., 12 (2011), 1062–1071. https://doi.org/10.1016/j.nonrwa.2010.08.028 doi: 10.1016/j.nonrwa.2010.08.028
|
| [9] |
R. Wu, Permanence of a discrete periodic volterra model with mutual interference and Beddington-Deangelis functional response, Discrete Dyn. Nat. Soc., 2010 (2010), 1038–1045. https://doi.org/10.1155/2010/246783 doi: 10.1155/2010/246783
|
| [10] |
E. Moustafa, M. A. Asmaa, Deterministic and stochastic fractional-order hastings-powell food chain model, Cmc-comput. Mater. Contin., 70 (2022), 2277–2296. https://doi.org/10.32604/cmc.2022.019314 doi: 10.32604/cmc.2022.019314
|
| [11] | B. Vahid, Y. M. Javad, T. M. Saleh, Global stabilization of Lotka-Volterra systems with interval uncertainty, 64 (2019), 1209–1213. https://doi.org/10.1109/TAC.2018.2845659 |
| [12] |
X. Zheng, D. Li, Interaction of Acidithiobacillus ferrooxidans, Rhizobium phaseoli and Rhodotorula sp. in bioleaching process based on Lotka-Volterra model, Electron. J. Biotechnol., 22 (2016), 90–97. https://doi.org/10.1016/j.ejbt.2016.06.004 doi: 10.1016/j.ejbt.2016.06.004
|
| [13] |
R. A. Cropp, J. Norbury, Population interactions in ecology: A rule-based approach to modeling ecosystems in a mass-conserving framework, SIAM Rev., 57 (2015), 437–465. https://doi.org/10.1137/140962528 doi: 10.1137/140962528
|
| [14] |
R. Arditi, H. Saiah, Empirical evidence of the role of heterogeneity in ratio-dependent consumption, Ecology, 73 (2008), 1544–1551. https://doi.org/10.2307/1940007 doi: 10.2307/1940007
|
| [15] |
X. Zhao, Z. Zeng, Stochastic dynamics of a two-species patch-system with ratio-dependent functional response, Qual. Theory Dyn. Syst., 21 (2022), 1–15. https://doi.org/10.1007/s12346-022-00594-x doi: 10.1007/s12346-022-00594-x
|
| [16] |
Q. Yang, X. Zhang, D. Jiang, M. Shao, Analysis of a stochastic predator-prey model with weak Allee effect and Holling-(n+1) functional response, Commun. Nonlinear Sci. Numer. Simul., 111 (2022), 106454. https://doi.org/10.1016/j.cnsns.2022.106454 doi: 10.1016/j.cnsns.2022.106454
|
| [17] |
P. A. Naik, Z. Eskandari, M. Yavuz, J. Zu, Complex dynamics of a discrete-time Bazykin-Berezovskaya prey-predator model with a strong Allee effect, J. Comput. Appl. Math., 413 (2022), 114401. https://doi.org/10.1016/j.cam.2022.114401 doi: 10.1016/j.cam.2022.114401
|
| [18] |
P. A. Naik, Z. Eskandari, Z. Avazzadeh, J. Zu, Multiple bifurcations of a discrete-time prey-predator model with mixed functional response, Int. J. Bifurcation Chaos, 32 (2022), 2250050. https://doi.org/10.1142/S021812742250050X doi: 10.1142/S021812742250050X
|
| [19] |
M. Hassell, Mutual interference between searching insect parasites, J. Anim. Ecol., 40 (1971), 473–486. https://doi.org/10.2307/3256 doi: 10.2307/3256
|
| [20] |
A. Watkinson, Density-dependence in single-species population, J. Anim. Ecol., 83 (1980), 345–357. https://doi.org/10.1016/0022-5193(80)90297-0 doi: 10.1016/0022-5193(80)90297-0
|
| [21] |
K. Wang, Y. Zhu, Global attractivity of positive periodic solution for a volterra model, Appl. Math. Comput., 203 (2008), 493–501. https://doi.org/10.1016/j.amc.2008.04.005 doi: 10.1016/j.amc.2008.04.005
|
| [22] | K. Wang, Stochastic biomathematics model, Sci. Press, 1 (2010), 1–207. |
| [23] |
K. Wang, Y. Zhu, Periodic solutions, permanence and global attractivity of a delayed impulsive prey-predator system with mutual interference, Nonlinear Anal. Real World Appl., 14 (2013). https://doi.org/10.1016/j.nonrwa.2012.08.016 doi: 10.1016/j.nonrwa.2012.08.016
|
| [24] |
K. Wang, Y. Zhu, Dynamics of a stochastic predator-prey model with mutual interference, Int. J. Biomath., 7 (2014), 1–22. https://doi.org/10.1142/S1793524514500260 doi: 10.1142/S1793524514500260
|
| [25] |
L. Wu, H. Zheng, S. Zhang, Dynamics of a non-autonomous predator-prey system with hassell-varley-hollingⅡfunction response and mutual interference, AIMS Math., 6 (2021), 6033–6049. https://doi.org/10.3934/math.2021355 doi: 10.3934/math.2021355
|
| [26] |
K. Gupta, S. Gakkhar, The filippov approach for predator-prey system involving mixed type of functional responses, Differ. Equations Dyn. Syst., 28 (2020), 1–21. https://doi.org/10.1007/s12591-016-0322-x doi: 10.1007/s12591-016-0322-x
|
| [27] |
X. Li, X. Lin, J. Liu, Existence and global attractivity of positive periodic solutions for a predator-prey model with crowley-martin functional response, Electron. J. Differ. Equations, 191 (2018), 1–17. https://doi.org/10.2298/FIL1901043L doi: 10.2298/FIL1901043L
|
| [28] |
L. Chen, F. Chen, Y. Wang, Influence of predator mutual interference and prey refuge on lotka-volterra predator-prey dynamics, Commun. Nonlinear Sci. Numer. Simul., 18 (2013), 3174–180. https://doi.org/10.1016/j.cnsns.2013.04.004 doi: 10.1016/j.cnsns.2013.04.004
|
| [29] |
Z. Ma, F. Chen, C. Wu, W. Chen, Dynamic behaviors of a lotka-volterra predator-prey model incorporating a prey refuge and predator mutual interference, Appl. Math. Comput., 219 (2013), 7945–7953. https://doi.org/10.1016/j.amc.2013.02.033 doi: 10.1016/j.amc.2013.02.033
|
| [30] |
C. Zhang, N. Huang, D. O'Regan, Almost periodic solutions for a Volterra model with mutual interference and Holling type Ⅲ functional response, Appl. Math. Comput., 225 (2013), 503–511. https://doi.org/10.1016/j.amc.2013.09.047 doi: 10.1016/j.amc.2013.09.047
|
| [31] |
X. Lin, F. Chen, Almost periodic solution for a Volterra model with mutual interference and Beddington-DeAngelis functional response, Appl. Math. Comput., 214 (2009), 548–556. https://doi.org/10.1016/j.amc.2009.04.028 doi: 10.1016/j.amc.2009.04.028
|
| [32] |
S. Arik, Global asymptotic stability analysis of bidirectional associative memory neural networks with time delays, IEEE Trans. Neural Networks, 16 (2003), 1–7. https://doi.org/10.1109/TNN.2005.844910 doi: 10.1109/TNN.2005.844910
|
| [33] |
B. Du, Existence, extinction and global asymptotical stability of a stochastic predator-prey model with mutual interference, J. Appl. Math. Comput., 46 (2014), 79–91. https://doi.org/10.1007/s12190-013-0738-1 doi: 10.1007/s12190-013-0738-1
|
| [34] |
D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. https://doi.org/10.1137/S0036144500378302 doi: 10.1137/S0036144500378302
|
Figures(1)
Pinglan Wan. Dynamic behavior of stochastic predator-prey system[J]. Electronic Research Archive, 2023, 31(5): 2925-2939. doi: 10.3934/era.2023147







 DownLoad:
DownLoad: