Cardiac arrhythmias are serious myocardial electrical disturbances that affect the rate and rhythm of heartbeats. Despite the rapidly accumulating data about the pathophysiology and the treatment, new insights are required to improve the overall clinical outcome of patients with cardiac arrhythmias. Three major arrhythmogenic processes can contribute to the pathogenesis of cardiac arrhythmias; 1) enhanced automaticity, 2) afterdepolarization-triggered activity and 3) reentry circuits. The mathematical model of the quantum tunneling of ions is used to investigate these mechanisms from a quantum mechanical perspective. The mathematical model focuses on applying the principle of quantum tunneling to sodium and potassium ions. This implies that these ions have a non-zero probability of passing through the gate, which has an energy that is higher than the kinetic energy of ions. Our mathematical findings indicate that, under pathological conditions, which affect ion channels, the quantum tunneling of sodium and potassium ions is augmented. This augmentation creates a state of hyperexcitability that can explain the enhanced automaticity, after depolarizations that are associated with triggered activity and a reentry circuit. Our mathematical findings stipulate that the augmented and thermally assisted quantum tunneling of sodium and potassium ions can depolarize the membrane potential and trigger spontaneous action potentials, which may explain the automaticity and afterdepolarization. Furthermore, the quantum tunneling of potassium ions during an action potential can provide a new insight regarding the formation of a reentry circuit. Introducing these quantum mechanical aspects may improve our understanding of the pathophysiological mechanisms of cardiac arrhythmias and, thus, contribute to finding more effective anti-arrhythmic drugs.
Citation: Mohammed I. A. Ismail, Abdallah Barjas Qaswal, Mo'ath Bani Ali, Anas Hamdan, Ahmad Alghrabli, Mohamad Harb, Dina Ibrahim, Mohammad Nayel Al-Jbour, Ibrahim Almobaiden, Khadija Alrowwad, Esra'a Jaibat, Mira Alrousan, Mohammad Banifawaz, Mohammed A. M. Aldrini, Aya Daikh, Nour Aldarawish, Ahmad Alabedallat, Ismail M. I. Ismail, Lou'i Al-Husinat. Quantum mechanical aspects of cardiac arrhythmias: A mathematical model and pathophysiological implications[J]. AIMS Biophysics, 2023, 10(3): 401-439. doi: 10.3934/biophy.2023024
Cardiac arrhythmias are serious myocardial electrical disturbances that affect the rate and rhythm of heartbeats. Despite the rapidly accumulating data about the pathophysiology and the treatment, new insights are required to improve the overall clinical outcome of patients with cardiac arrhythmias. Three major arrhythmogenic processes can contribute to the pathogenesis of cardiac arrhythmias; 1) enhanced automaticity, 2) afterdepolarization-triggered activity and 3) reentry circuits. The mathematical model of the quantum tunneling of ions is used to investigate these mechanisms from a quantum mechanical perspective. The mathematical model focuses on applying the principle of quantum tunneling to sodium and potassium ions. This implies that these ions have a non-zero probability of passing through the gate, which has an energy that is higher than the kinetic energy of ions. Our mathematical findings indicate that, under pathological conditions, which affect ion channels, the quantum tunneling of sodium and potassium ions is augmented. This augmentation creates a state of hyperexcitability that can explain the enhanced automaticity, after depolarizations that are associated with triggered activity and a reentry circuit. Our mathematical findings stipulate that the augmented and thermally assisted quantum tunneling of sodium and potassium ions can depolarize the membrane potential and trigger spontaneous action potentials, which may explain the automaticity and afterdepolarization. Furthermore, the quantum tunneling of potassium ions during an action potential can provide a new insight regarding the formation of a reentry circuit. Introducing these quantum mechanical aspects may improve our understanding of the pathophysiological mechanisms of cardiac arrhythmias and, thus, contribute to finding more effective anti-arrhythmic drugs.

| [1] |
Huikuri HV, Castellanos A, Myerburg RJ (2001) Sudden death due to cardiac arrhythmias. New Engl J Med 345: 1473-1482. https://doi.org/10.1056/NEJMra000650 
|
| [2] |
Srinivasan NT, Schilling RJ (2018) Sudden cardiac death and arrhythmias. Arrhyth Electrophysi Rev 7: 111. https://doi.org/10.15420/aer.2018:15:2
|
| [3] |
John RM, Tedrow UB, Koplan BA, et al. (2012) Ventricular arrhythmias and sudden cardiac death. Lancet 380: 1520-1529. https://doi.org/10.1016/S0140-6736(12)61413-5
|
| [4] |
Janse MJ, Wit AL (1989) Electrophysiological mechanisms of ventricular arrhythmias resulting from myocardial ischemia and infarction. Physiol Rev 69: 1049-1169. https://doi.org/10.1152/physrev.1989.69.4.1049
|
| [5] |
Peretto G, Sala S, Rizzo S, et al. (2019) Arrhythmias in myocarditis: state of the art. Heart Rhythm 16: 793-801. https://doi.org/10.1016/j.hrthm.2018.11.024
|
| [6] |
Kumar S, Stevenson WG, John RM (2015) Arrhythmias in dilated cardiomyopathy. Card Electrophy Clin 7: 221-233. https://doi.org/10.1016/j.hrthm.2018.11.024
|
| [7] |
Tisdale JE, Chung MK, Campbell KB, et al. (2020) Drug-induced arrhythmias: a scientific statement from the American Heart Association. Circulation 142: e214-233. https://doi.org/10.1161/CIR.0000000000000905
|
| [8] |
Behere SP, Weindling SN (2015) Inherited arrhythmias: The cardiac channelopathies. Ann Pediat Cardiol 8: 210. https://doi.org/10.4103/0974-2069.164695
|
| [9] |
FISCH C (1973) Relation of electrolyte disturbances to cardiac arrhythmias. Circulation 47: 408-419. https://doi.org/10.1161/01.CIR.47.2.408
|
| [10] |
Tse G (2016) Mechanisms of cardiac arrhythmias. J Arrhythm 32: 75-81. https://doi.org/10.1016/j.joa.2015.11.003
|
| [11] |
Antzelevitch C, Burashnikov A (2011) Overview of basic mechanisms of cardiac arrhythmia. Card Electrophy Clin 3: 23-45. https://doi.org/10.1016/j.ccep.2010.10.012
|
| [12] |
Marbán E (2002) Cardiac channelopathies. Nature 415: 213-218. https://doi.org/10.1038/415213a
|
| [13] |
Franz MR, Cima R, Wang D, et al. (1992) Electrophysiological effects of myocardial stretch and mechanical determinants of stretch-activated arrhythmias. Circulation 86: 968-978. https://doi.org/10.1161/01.CIR.86.3.968
|
| [14] | Morand J, Arnaud C, Pepin JL, et al. (2018) Chronic intermittent hypoxia promotes myocardial ischemia-related ventricular arrhythmias and sudden cardiac death. Sci Rep 8: 1-8. https://doi.org/10.1038/s41598-018-21064-y |
| [15] |
Orchard CH, Cingolani HE (1994) Acidosis and arrhythmias in cardiac muscle. Card Res 28: 1312-1319. https://doi.org/10.1093/cvr/28.9.1312
|
| [16] |
Morris CE (2011) Voltage-gated channel mechanosensitivity: fact or friction?. Front Physiol 2: 25. https://doi.org/10.3389/fphys.2011.00025
|
| [17] | Dehghani-Samani A, Madreseh-Ghahfarokhi S, Dehghani-Samani A (2019) Mutations of voltage-gated ionic channels and risk of severe cardiac arrhythmias. Acta Cardiol Sin 35: 99. https://doi.org/10.6515%2FACS.201903_35(2).20181028A |
| [18] |
Li Q, Huang H, Liu G, et al. (2009) Gain-of-function mutation of Nav1. 5 in atrial fibrillation enhances cellular excitability and lowers the threshold for action potential firing. Biochem Biophys Res Commun 380: 132-137. https://doi.org/10.1016/j.bbrc.2009.01.052
|
| [19] | Moskalenko A (2014) Cardiac Arrhythmias Mechanisms, Pathophysiology, and Treatment: 1-162. https://doi.org/10.5772/57008 |
| [20] | Cardiac Arrhythmia Suppression Trial (CAST) Investigators.Preliminary report: effect of encainide and flecainide on mortality in a randomized trial of arrhythmia suppression after myocardial infarction. N Engl J Med (1989) 321: 406-412. https://doi.org/10.1056/nejm198908103210629 |
| [21] |
Brooks MM, Gorkin L, Schron EB, et al. (1994) Moricizine and quality of life in the Cardiac Arrhythmia Suppression Trial II (CAST II). Control Clin Trials 15: 437-449. https://doi.org/10.1016/0197-2456(94)90002-7
|
| [22] |
Kurian TK, Efimov IR (2010) Mechanisms of fibrillation: Neurogenic or myogenic? reentrant or focal? multiple or single?: Still puzzling after 160 years of inquiry. J Card Electrophysiol 21: 1274. https://doi.org/10.1111%2Fj.1540-8167.2010.01820.x
|
| [23] |
Calvillo L, Redaelli V, Ludwig N, et al. (2022) Quantum biology research meets pathophysiology and therapeutic mechanisms: a biomedical perspective. Quantum Rep 4: 148-172. https://www.mdpi.com/2624-960X/4/2/11
|
| [24] |
Kim Y, Bertagna F, D'souza EM, et al. (2021) Quantum biology: An update and perspective. Quantum Rep 3: 80-126. https://www.mdpi.com/2624-960X/3/1/6
|
| [25] |
Cao J, Cogdell RJ, Coker DF, et al. (2020) Quantum biology revisited. Sci Adv 6: eaaz4888. https://doi.org/10.1126/sciadv.aaz4888
|
| [26] |
Slocombe L, Sacchi M, Al-Khalili (2022) An open quantum systems approach to proton tunnelling in DNA. Commun Phys 5: 109. https://doi.org/10.1038/s42005-022-00881-8
|
| [27] |
Sutcliffe MJ, Scrutton NS (2002) A new conceptual framework for enzyme catalysis: Hydrogen tunneling coupled to enzyme dynamics in flavoprotein and quinoprotein enzymes. Eur J Biochem 269: 3096-3102. https://doi.org/10.1046/j.1432-1033.2002.03020.x
|
| [28] |
Qaswal AB (2019) Quantum tunneling of ions through the closed voltage-gated channels of the biological membrane: A mathematical model and implications. Quantum Rep 1: 219-225. https://www.mdpi.com/2624-960X/1/2/19
|
| [29] | Miller DA (2008). Quantum mechanics for scientists and engineers. Cambridge University Press |
| [30] |
Aryal P, Sansom MS, Tucker SJ (2015) Hydrophobic gating in ion channels. J Mol Biol 427: 121-130. https://doi.org/10.1016/j.jmb.2014.07.030
|
| [31] |
Oelstrom K, Goldschen-Ohm MP, Holmgren M, et al. (2014) Evolutionarily conserved intracellular gate of voltage-dependent sodium channels. Nat Commun 5: 3420. https://doi.org/10.1038/ncomms4420
|
| [32] |
Jensen MØ, Borhani DW, Lindorff-Larsen K, et al. (2010) Principles of conduction and hydrophobic gating in K+ channels. Proceedings of the National Academy of Sciences 107: 5833-5838. https://doi.org/10.1073/pnas.0911691107
|
| [33] |
Trick JL, Aryal P, Tucker SJ, et al. (2015) Molecular simulation studies of hydrophobic gating in nanopores and ion channels. Biochem Society Transact 43: 146-150. https://doi.org/10.1042/BST20140256
|
| [34] |
Rao S, Klesse G, Lynch CI, et al. (2021) Molecular simulations of hydrophobic gating of pentameric ligand gated ion channels: insights into water and ions. J Phys Chem B 125: 981-994. https://doi.org/10.1021/acs.jpcb.0c09285
|
| [35] |
Chandra AK (1974). |
| [36] |
Miyazaki T (2004). |
| [37] | Serway RA, Moses CJ, Moyer CA (2004). Modern physics |
| [38] |
Eckart C (1930) The penetration of a potential barrier by electrons. Phys Rev 35: 1303. https://doi.org/10.1103/PhysRev.35.1303
|
| [39] |
Zhu F, Hummer G (2012) Drying transition in the hydrophobic gate of the GLIC channel blocks ion conduction. Biophys J 103: 219-227. http://dx.doi.org/10.1016/j.bpj.2012.06.003
|
| [40] |
Rao S, Lynch CI, Klesse G, et al. (2018) Water and hydrophobic gates in ion channels and nanopores. Faraday Discuss 209: 231-247. https://doi.org/10.1039/C8FD00013A
|
| [41] |
Neale C, Chakrabarti N, Pomorski P, et al. (2015) Hydrophobic gating of ion permeation in magnesium channel CorA. PLoS Comput Biol 11: e1004303. https://doi.org/10.1371/journal.pcbi.1004303
|
| [42] |
Khavrutskii IV, Gorfe AA, Lu B, et al. (2009) Free energy for the permeation of Na+ and Cl− ions and their ion-pair through a zwitterionic dimyristoyl phosphatidylcholine lipid bilayer by umbrella integration with harmonic fourier beads. J Am Chem Society 131: 1706-1716. https://doi.org/10.1021/ja8081704
|
| [43] |
Vorobyov I, Olson TE, Kim JH, et al. (2014) Ion-induced defect permeation of lipid membranes. Biophys J 106: 586-597. http://dx.doi.org/10.1016/j.bpj.2013.12.027
|
| [44] |
Leontiadou H, Mark AE, Marrink SJ (2007) Ion transport across transmembrane pores. Biophys J 92: 4209-4215. http://dx.doi.org/10.1529/biophysj.106.101295
|
| [45] |
Zhang HY, Xu Q, Wang YK, et al. (2016) Passive transmembrane permeation mechanisms of monovalent ions explored by molecular dynamics simulations. J Chem Theory Comput 12: 4959-4969. https://doi.org/10.1021/acs.jctc.6b00695
|
| [46] |
Chen F, Hihath J, Huang Z, et al. (2007) Measurement of single-molecule conductance. Annu Rev Phys Chem 58: 535-564. https://doi.org/10.1146/annurev.physchem.58.032806.104523
|
| [47] |
Qaswal AB (2020) Quantum electrochemical equilibrium: Quantum version of the Goldman–Hodgkin–Katz equation. Quantum Rep 2: 266-277. https://www.mdpi.com/2624-960X/2/2/17
|
| [48] |
Qaswal AB (2021) The role of quantum tunneling of ions in the pathogenesis of the cardiac arrhythmias due to channelopathies, ischemia, and mechanical stretch. Biophysics 66: 637-641. https://doi.org/10.1134/S0006350921040072
|
| [49] |
Ababneh O, Qaswal AB, Alelaumi A, et al. (2021) Proton quantum tunneling: Influence and relevance to acidosis-induced cardiac arrhythmias/cardiac arrest. Pathophysiology 28: 400-436. https://www.mdpi.com/1873-149X/28/3/27
|
| [50] |
Zhang XC, Yang H, Liu Z, et al. (2018) Thermodynamics of voltage-gated ion channels. Biophys Rep 4: 300-319. https://doi.org/10.1016/j.celrep.2021.109931
|
| [51] |
Summhammer J, Salari V, Bernroider G (2012) A quantum-mechanical description of ion motion within the confining potentials of voltage-gated ion channels. J Integr Neurosci 11: 123-135. https://doi.org/10.1142/S0219635212500094
|
| [52] |
Summhammer J, Sulyok G, Bernroider G (2018) Quantum dynamics and non-local effects behind ion transition states during permeation in membrane channel proteins. Entropy 20: 558. https://doi.org/10.1142/S0219635212500094
|
| [53] |
Summhammer J, Sulyok G, Bernroider G (2020) Quantum mechanical coherence of K+ ion wave packets increases conduction in the KcsA ion channel. Appl Sci 10: 4250. https://www.mdpi.com/2076-3417/10/12/4250
|
| [54] |
Wang K, Wang S, Yang L, et al. (2021) THz trapped ion model and THz spectroscopy detection of potassium channels. Nano Res 15: 3825-3833. https://doi.org/10.1007/s12274-021-3965-z
|
| [55] |
Karandashev K, Xu ZH, Meuwly M, et al. (2017) Kinetic isotope effects and how to describe them. Struct Dynam 4: 061501. https://doi.org/10.1063/1.4996339
|
| [56] |
Sen A, Kohen A (2010) Enzymatic tunneling and kinetic isotope effects: chemistry at the crossroads. J Phys Org Chem 23: 613-619. https://doi.org/10.1002/poc.1633
|
| [57] |
Eckhardt AK, Gerbig D, Schreiner PR (2018) Heavy atom secondary kinetic isotope effect on H-tunneling. J Phys Chem A 122: 1488-1495. https://doi.org/10.1021/acs.jpca.7b12118
|
| [58] |
Nappi P, Miceli F, Soldovieri MV, et al. (2020) Epileptic channelopathies caused by neuronal Kv7 (KCNQ) channel dysfunction. Pflüg Arch-Eur J Phy 472: 881-898. https://doi.org/10.1007/s00424-020-02404-2
|
| [59] | Niday Z, Tzingounis AV (2018) Potassium channel gain of function in epilepsy: an unresolved paradox. Neurosci 24: 368-380. https://doi.org/10.1177/1073858418763752 |
| [60] |
Miceli F, Soldovieri MV, Ambrosino P, et al. (2015) Early-onset epileptic encephalopathy caused by gain-of-function mutations in the voltage sensor of Kv7. 2 and Kv7. 3 potassium channel subunits. J Neurosci 35: 3782-3793. https://doi.org/10.1523/JNEUROSCI.4423-14.2015
|
| [61] |
Du W, Bautista JF, Yang H, et al. (2005) Calcium-sensitive potassium channelopathy in human epilepsy and paroxysmal movement disorder. Nat Genet 37: 733-738. https://doi.org/10.1038/ng1585
|
| [62] |
Robinson RB, Siegelbaum SA (2003) Hyperpolarization-activated cation currents: from molecules to physiological function. Annu Rev Physiol 65: 453-480. https://doi.org/10.1146/annurev.physiol.65.092101.142734
|
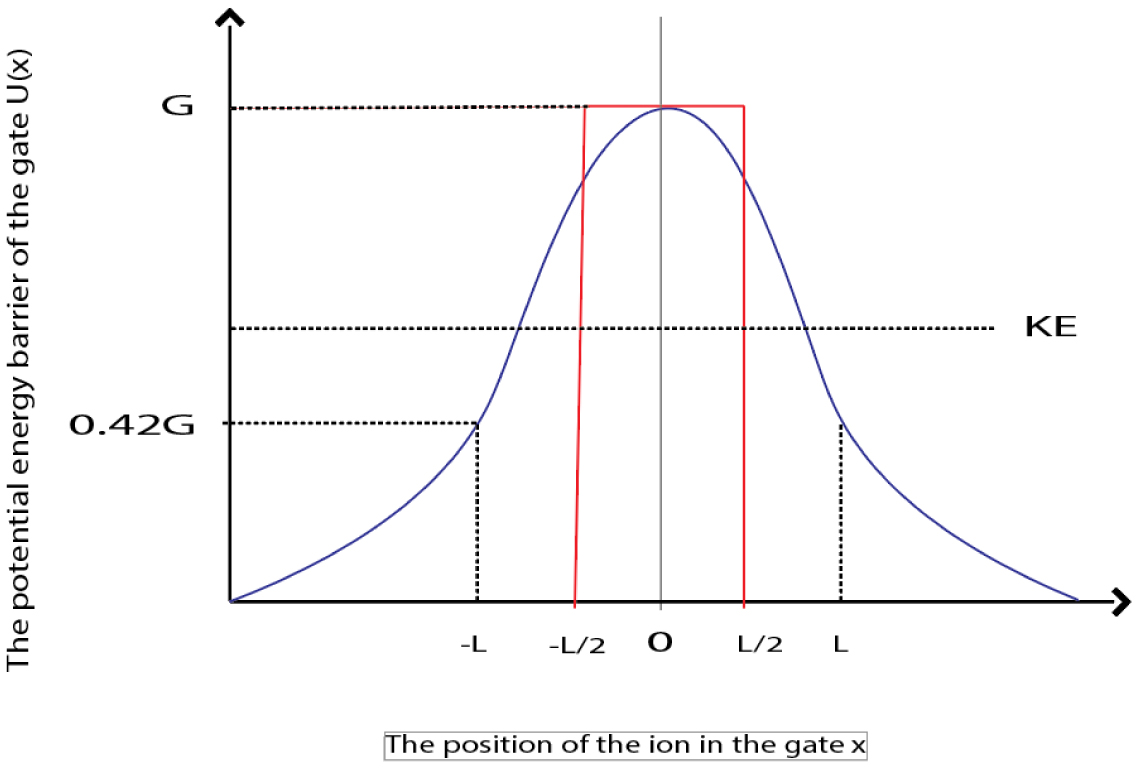



















Figures(21) / Tables(1)

Mohammed I. A. Ismail, Abdallah Barjas Qaswal, Mo'ath Bani Ali, Anas Hamdan, Ahmad Alghrabli, Mohamad Harb, Dina Ibrahim, Mohammad Nayel Al-Jbour, Ibrahim Almobaiden, Khadija Alrowwad, Esra'a Jaibat, Mira Alrousan, Mohammad Banifawaz, Mohammed A. M. Aldrini, Aya Daikh, Nour Aldarawish, Ahmad Alabedallat, Ismail M. I. Ismail, Lou'i Al-Husinat. Quantum mechanical aspects of cardiac arrhythmias: A mathematical model and pathophysiological implications[J]. AIMS Biophysics, 2023, 10(3): 401-439. doi: 10.3934/biophy.2023024







 DownLoad:
DownLoad: