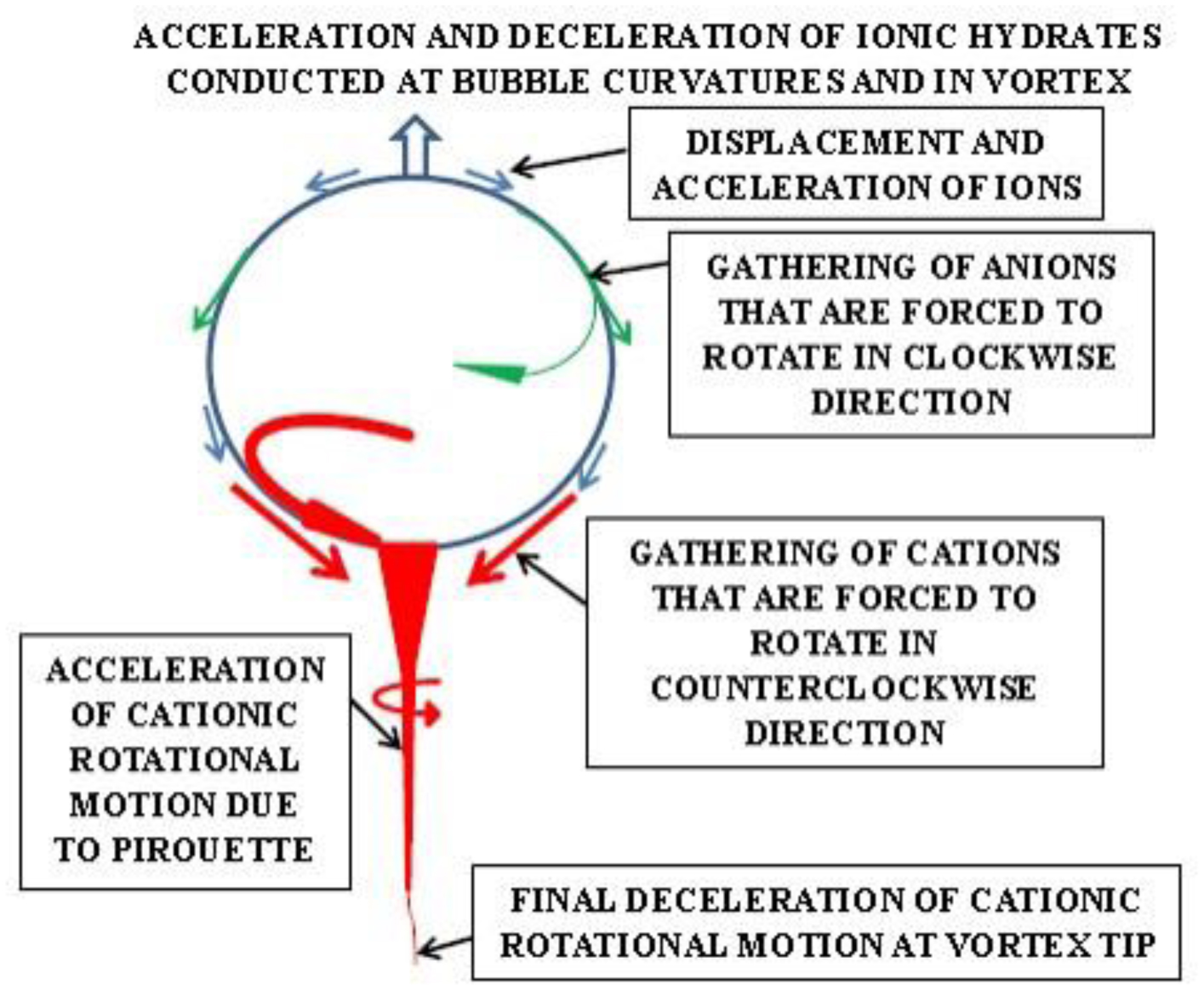
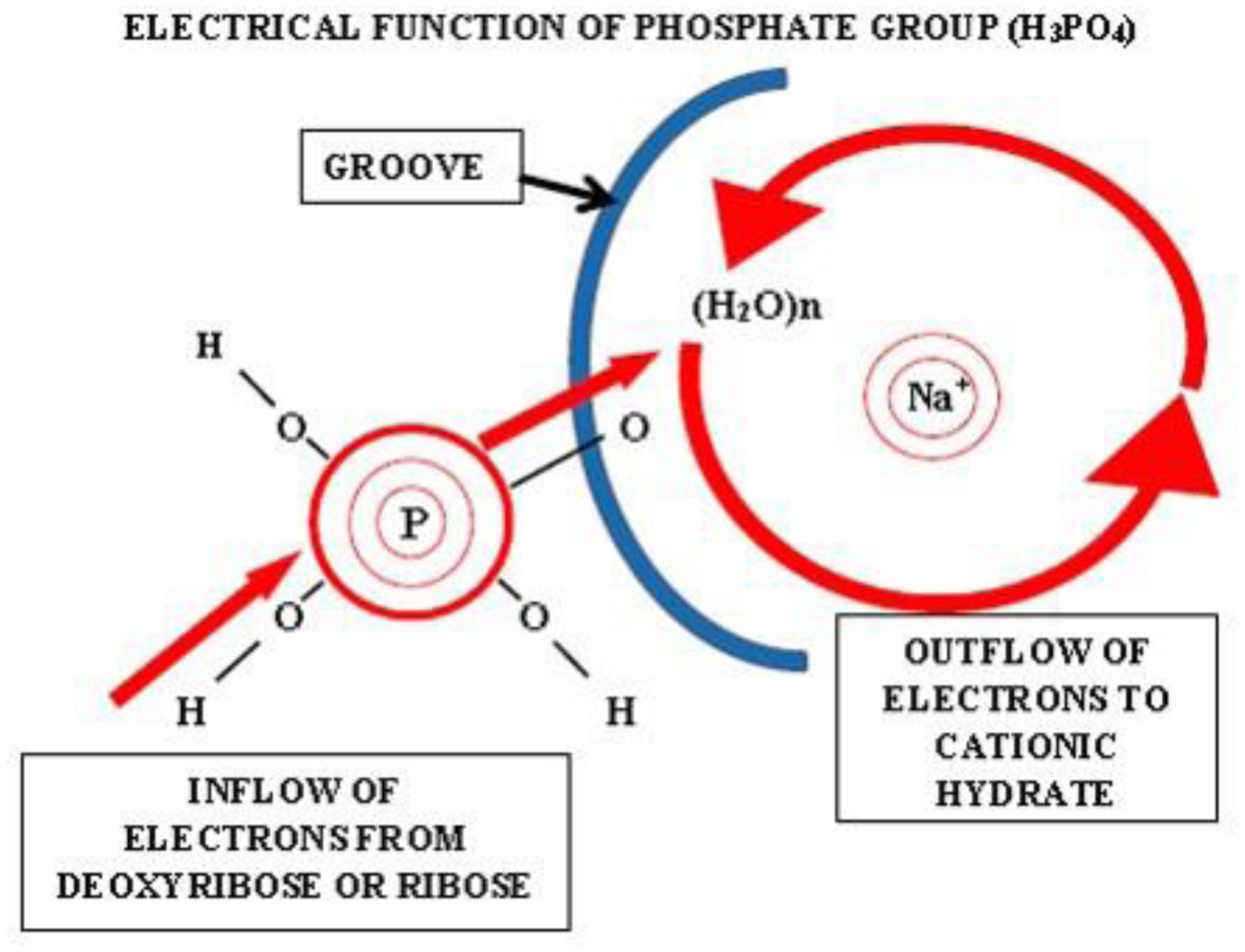
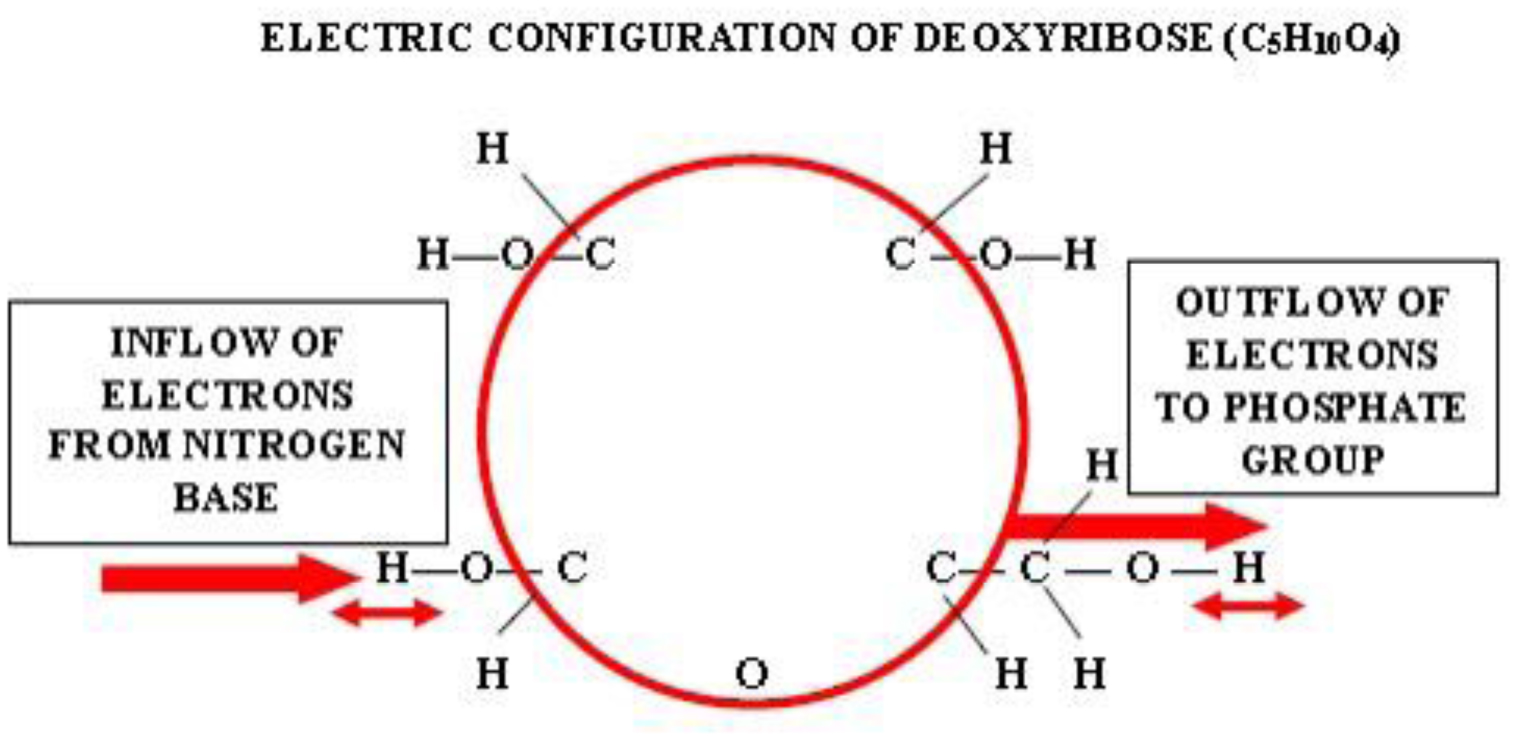
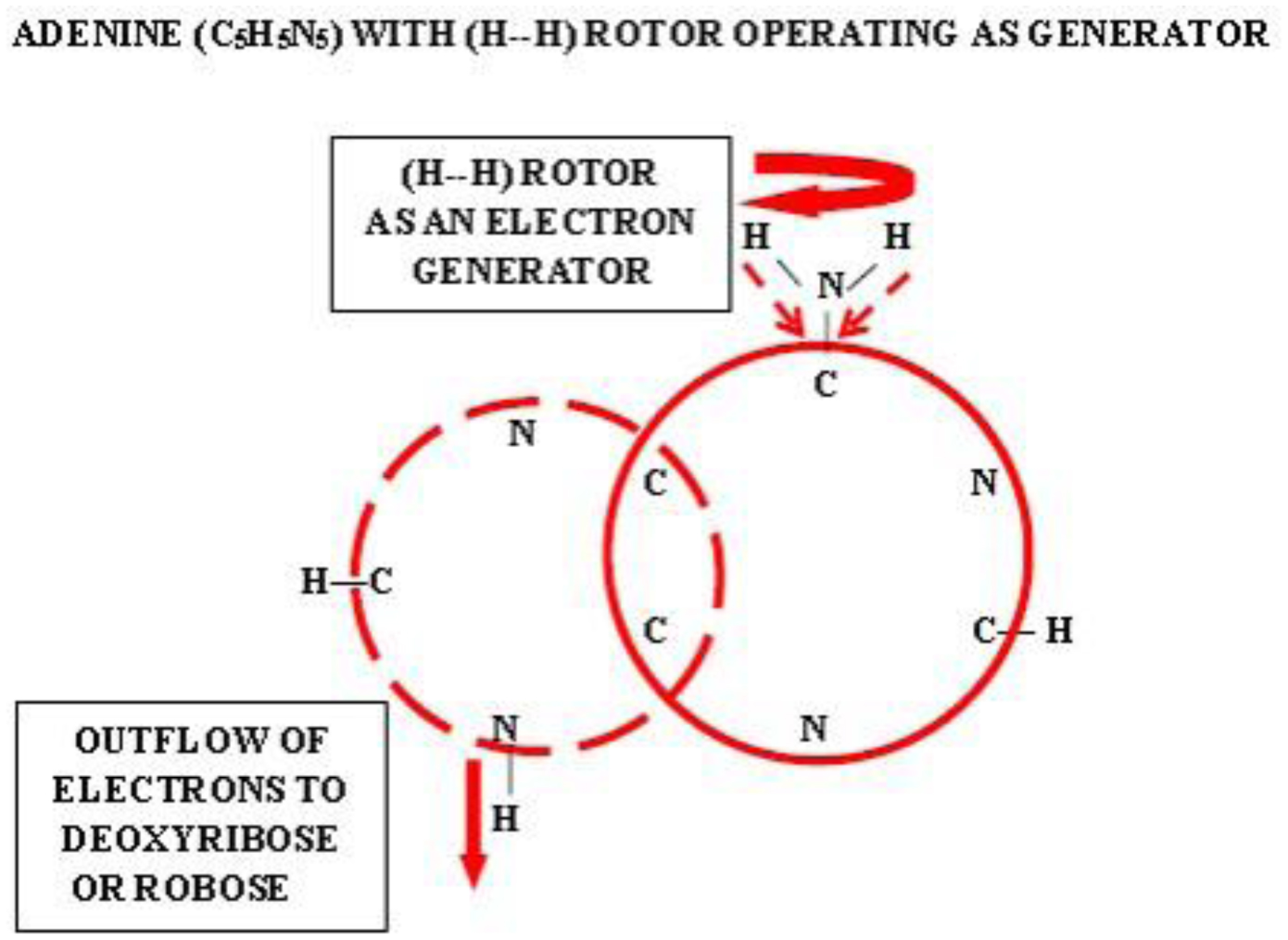
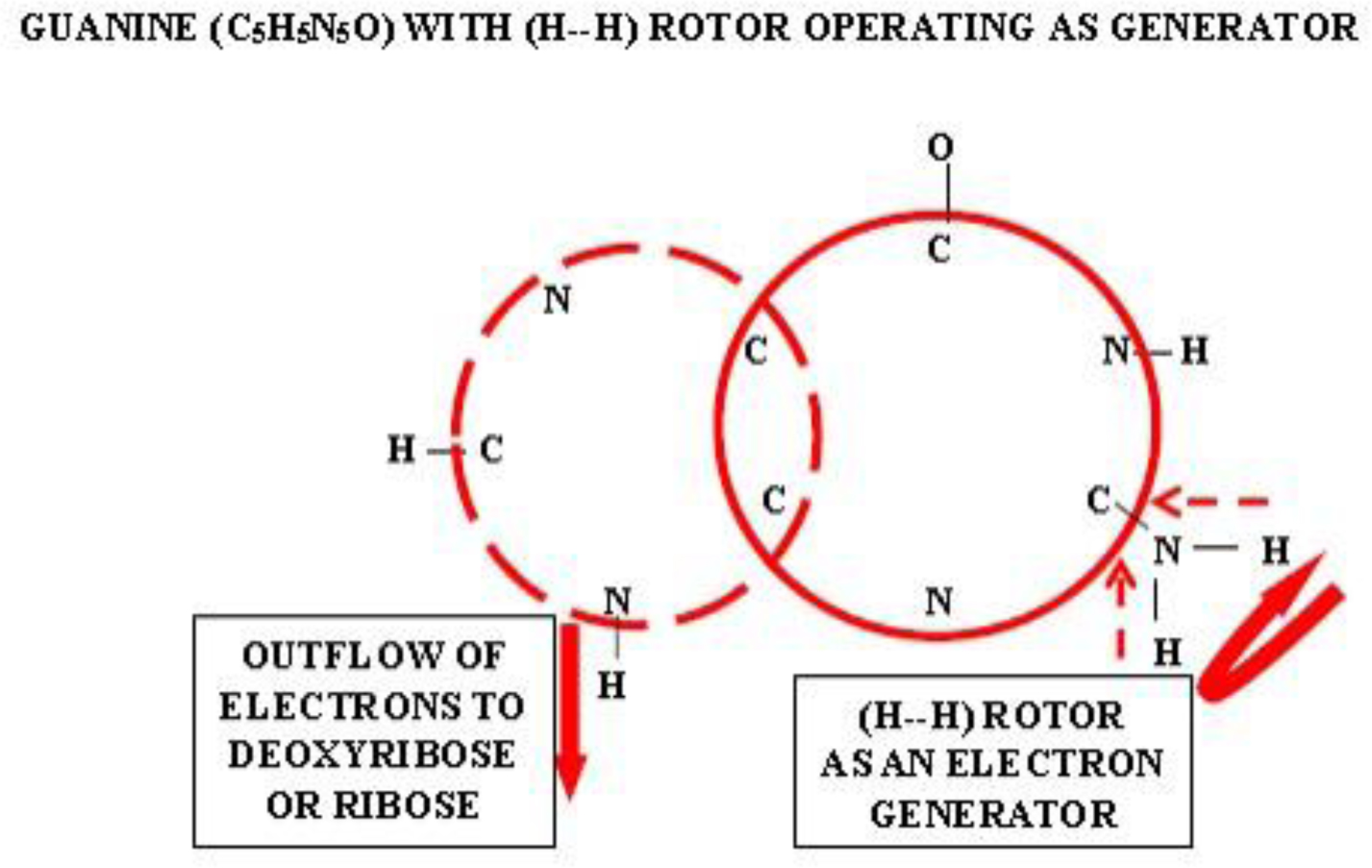
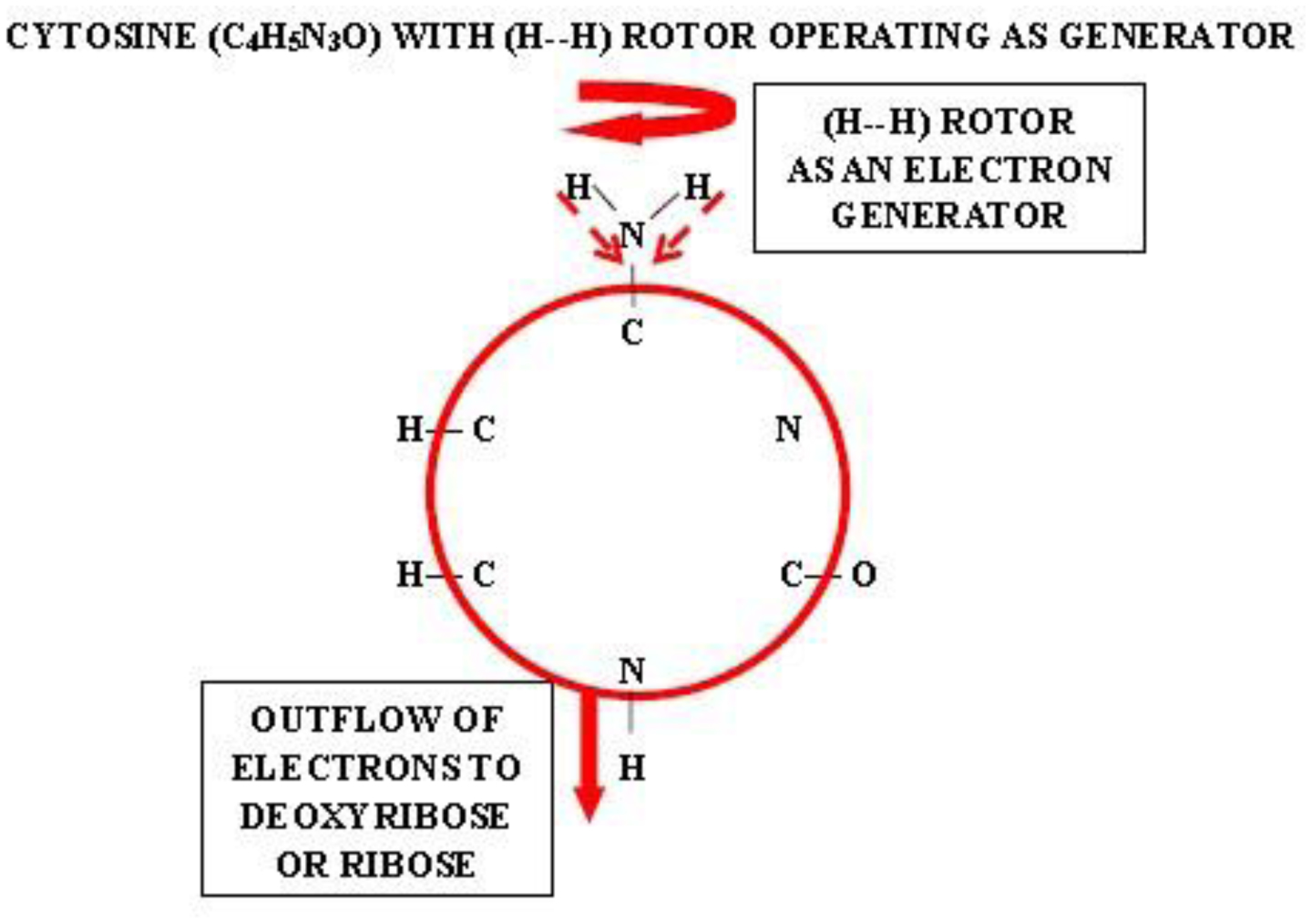
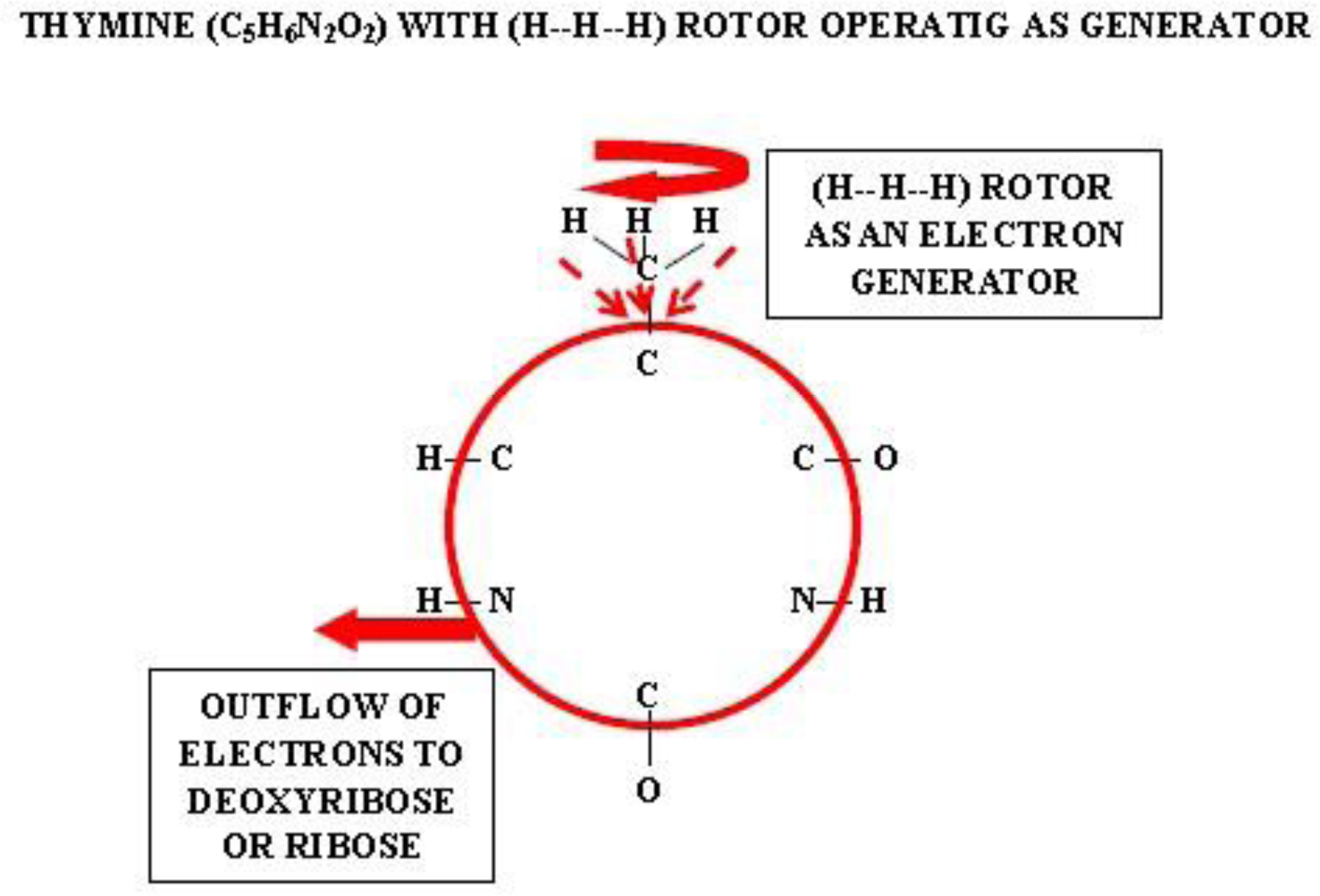
Photographic investigations of rising bubbles in seawater revealed that each bubble may conduct a single or bi-spiraling motion, which resemble architecture of RNA or DNA respectively. The rotational motion results from acceleration of ionic hydrates, which are separated to anionic and cationic domains at the upper and bottom curvatures of the bubble. Afterwards, rotational motion undergoes further acceleration in the bubble upper vortex, followed by deceleration at the vortex tip. During that phase, the spiraling motion cause significant friction that result in polarization of electronegative atoms of H, C, N, O and P. These may be simultaneously arranged around a whirling cationic strands and form phosphate groups, ribose and nitrogen bases equipped with H2 and H3 rotors. It is hypothesized that such hydrogen rotors may operate as generators of electrons, which may be detached from valence shells of electropositive atoms. Then, electrons may flow via nitrogen bases and deoxyribose or ribose to phosphate groups. Next, the negatively charged edges of phosphate groups may attract cationic hydrates and energize their rotational motion in the grooves, then causing also its spiraling projection outward. That may be responsible for replication of nucleotides and its arrangement along the cationic flow into RNA or DNA polymers, in the same manner as originally produced by rising bubbles. Moreover, it points that hydrogen rotors may generate energy needed for viability as well as interact with all physical and chemical fields.
Citation: Roman Marks, Piotr H. Pawłowski. Rotational-electric principles of RNA/DNA and viability[J]. AIMS Biophysics, 2023, 10(3): 385-400. doi: 10.3934/biophy.2023023
Photographic investigations of rising bubbles in seawater revealed that each bubble may conduct a single or bi-spiraling motion, which resemble architecture of RNA or DNA respectively. The rotational motion results from acceleration of ionic hydrates, which are separated to anionic and cationic domains at the upper and bottom curvatures of the bubble. Afterwards, rotational motion undergoes further acceleration in the bubble upper vortex, followed by deceleration at the vortex tip. During that phase, the spiraling motion cause significant friction that result in polarization of electronegative atoms of H, C, N, O and P. These may be simultaneously arranged around a whirling cationic strands and form phosphate groups, ribose and nitrogen bases equipped with H2 and H3 rotors. It is hypothesized that such hydrogen rotors may operate as generators of electrons, which may be detached from valence shells of electropositive atoms. Then, electrons may flow via nitrogen bases and deoxyribose or ribose to phosphate groups. Next, the negatively charged edges of phosphate groups may attract cationic hydrates and energize their rotational motion in the grooves, then causing also its spiraling projection outward. That may be responsible for replication of nucleotides and its arrangement along the cationic flow into RNA or DNA polymers, in the same manner as originally produced by rising bubbles. Moreover, it points that hydrogen rotors may generate energy needed for viability as well as interact with all physical and chemical fields.

| [1] | Blanchard DC, Woodcock AH (1957) Bubble formation and modification in the sea and its meteorological significance. Tellus 9: 145-158. https://doi.org/10.3402/tellusa.v9i2.9094 |
| [2] |
Monahan EC (1971) Oceanic whitecaps. J Phys Oceanogr 1: 139-144. https://doi.org/10.1175/1520-0485(1971)001%3C0139:OW%3E2.0.CO;2 
|
| [3] |
Liss PS (1983) Gas transfer: experiments and geochemical implications. Air-Sea Exchange of Gases and Particles . Dordrecht: Springer Netherlands 241-298. https://doi.org/10.1007/978-94-009-7169-1_5
|
| [4] | Garbalewski C, Marks R (1987) Latitudinal characteristics of aerosol distribution in the near surface air over the Atlantic. Acta Geophys Pol 35: 77-86. |
| [5] |
Stramska M, Marks R, Monahan EC (1990) Bubble-mediated aerosol production as a consequence of wave breaking in supersaturated (hyperoxic) seawater. J Geophys Res Oceans 95: 18281-18288. https://doi.org/10.1029/JC095iC10p18281
|
| [6] |
Woolf DK, Thorpe SA (1991) Bubbles and the air-sea exchange of gases in near saturation conditions. J Mar Res 49: 435-466.
|
| [7] | Marks R (2015) Sub-bubble bi-pirouette splicing of cationic and anionic bases as a process of RNA/DNA creation. Oceanography 2: 128. https://www.researchgate.net/publication/284766375 |
| [8] |
Marks R (2022) Bubble mediated polymerization of RNA and DNA. AIMS Biophys 9: 96-107. https://doi.org/10.3934/biophy.2022009
|
| [9] |
Marks R (2019) Water vpor iduced arborne rtational fatures. Meteorol Hydrol Water Manage 7: 39-47. https://doi.org/10.26491/mhwm/104634
|
| [10] | Marks R (2014) Bubble rotational features-preliminary investigations. Oceanography 2: 128. https://doi.org/10.4172/2332-2632.1000128 |
| [11] |
Marks R, Górecka E, McCartney K, et al. (2019) Rising bubbles as mechanism for scavenging and aerosolization of diatoms. J Aerosol Sci 128: 79-88. https://doi.org/10.1016/j.jaerosci.2018.12.003
|
| [12] | Larmor J (1987) On the theory of the magnetic influence on spectra; and on the radiation from moving ions. Philosoph Magazine 44: 503-512. https://doi:10.1080/14786449708621095 |
| [13] |
Lipfert J, Doniach S, Das R, et al. (2014) Understanding of nucleic acid-ion interactions. Annu Rev Biochem 83: 813-841. https://doi.org/10.1146/annurev-biochem-060409-092720
|
| [14] |
Slater JC (1964) Atomic Radii in crystals. J Chem Phys 41: 3199-3204. https://doi.org/10.1063/1.1725697
|
| [15] |
Miller SL, Urey HC (1959) Organic compound synthesis on the primitive earth. Science 130: 245-251. https://doi.org/10.1126/science.130.3370.245
|
| [16] |
Benner SA, Kim HJ, Carrigan MA (2012) Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Acc Chem Res 45: 2025-2034. https://doi.org/10.1021/ar200332w
|
| [17] | Pawłowski PH (2019) The smooth evolution of the universal genetic code. Main episodes. Inter J Sci 8: 28-51. https://doi.org/10.18483/ijSci.218019 |
| [18] |
de Farias ST, dos Santos Junior AP, Rêgo TG, et al. (2017) Origin and evolution of RNA-dependent RNA polymerase. Front Genet 8: 125. https://doi.org/10.3389/fgene.2017.00125
|
| [19] |
Draper DE (2008) RNA folding: Thermodynamic and molecular descriptions of the role of ions. Biophys J 95: 5489-5495. https://doi.org/10.1529/biophysj.108.131813
|
| [20] | Eisenberg R (2023) Kirchhoff coupling generates ATP, the chemical energy of life. Qeios . https://doi.org/10.32388/C8DYYK |
| [21] |
Blanchard DC, Syzdek LD (1970) Mechanism for the water-to-air transfer and concentration of bacteria. Science 170: 626-628.
|
| [22] |
Marks R, Kruczalak K, Jankowska K, et al. (2001) Bacteria and Fungi in air over the Gulf of Gdańsk and Baltic Sea. J Aerosol Sci 32: 43-56. https://doi.org/10.1016/S0021-8502(00)00064-1
|
| [23] |
Aller JY, Kuznetsova MR, Jahns C, et al. (2005) The sea surface microlayer as a source of viral and bacterial enrichment in marine aerosol. J Aerosol Sci 36: 801-812. https://doi/10.1016/j.jaerosci.2004.10.012
|
| [24] |
Michaud JM, Thompson LR, Kaul D, et al. (2018) Taxon-specific aerosolization of bacteria and viruses in an experimental ocean-atmosphere mesocosm. Nat Commun 9: 2017. https://doi.org/10.1038/s41467-018-04409-z
|
| [25] |
Reche I. D'Orta G, D'Orta G, Mladenov N, et al. (2018) Deposition rates of viruses and bacteria above the atmospheric boundary layer. ISME J 12: 1154-1162. https://doi.org/10.1038/s41396-017-0042-4
|
| [26] |
Pawłowski PH (2021) Charged amino acids may promote coronavirus SARS-CoV-2 fusion with the host cell. AIMS Biophys 8: 111-120. https://doi.org/10.3934/biophys.2021008
|
| [27] | Xu K (2013) DNA circuit system and charge transfer mechanism. Engineering 5: 382-385. https://doi.org/10.4236/eng.2013.510B077 |
| [28] | Roberts GCK (2013) UV absorbance spectroscopy of biological macromolecules. Encyclopedia of Biophysics : 1-5. https://doi.org/10.1007/978-3-642-16712-6_780 |
| [29] |
Chou PJ, Johnson WC (1993) Base inclinations in natural and synthetic DNAs. J Am Chem Soc 115: 1205-1214. https://doi.org/10.1021/ja00057a001
|
| [30] |
Cherstvy AG, Kornyshev AA (2005) DNA melting in aggregates: Impeded or facilitated?. J Phys Chem B 109: 13024-13029. https://doi.org/10.1021/jp051117i
|
| [31] |
Cherstvy AG (2005) Structure of DNA toroids and electrostatic attraction of DNA duplexes. J Phys Condens Matter 17: 1363. https://doi.org/10.1088/0953-8984/17/8/015
|
| [32] |
Baldwin GS, Brooks NJ, Robson RE, et al. (2008) DNA double helices recognize mutual sequence homology in a protein free environment. J Phys Chem B 112: 1060-1064. https://doi.org/10.1021/jp7112297
|
| [33] |
Kornyshev AA, Lee DJ, Wynveen A (2017) Theory of phase segregation in DNA assemblies containing two different base-pair sequence types. New J Phys 19: 015014. https://doi.org/10.1088/1367-2630/aa5482
|
| [34] |
Dominic J (2018) Introducing a model of pairing based on base pair specific interactions between identical DNA sequences. J Phys Condens Matter 30: 075102. https://doi.org/10.1088/1361-648X/aaa043
|
| [35] |
Gladyshev E, Kleckner N (2018) Recombination-independent recognition of DNA homology for repeat-induced point mutation (RIP) is modulated by the underlying nucleotide sequence. PLoS Genet 12: e1006015. https://doi.org/10.1371/journal.pgen.1006015
|
Figures(7)

Roman Marks, Piotr H. Pawłowski. Rotational-electric principles of RNA/DNA and viability[J]. AIMS Biophysics, 2023, 10(3): 385-400. doi: 10.3934/biophy.2023023







 DownLoad:
DownLoad: