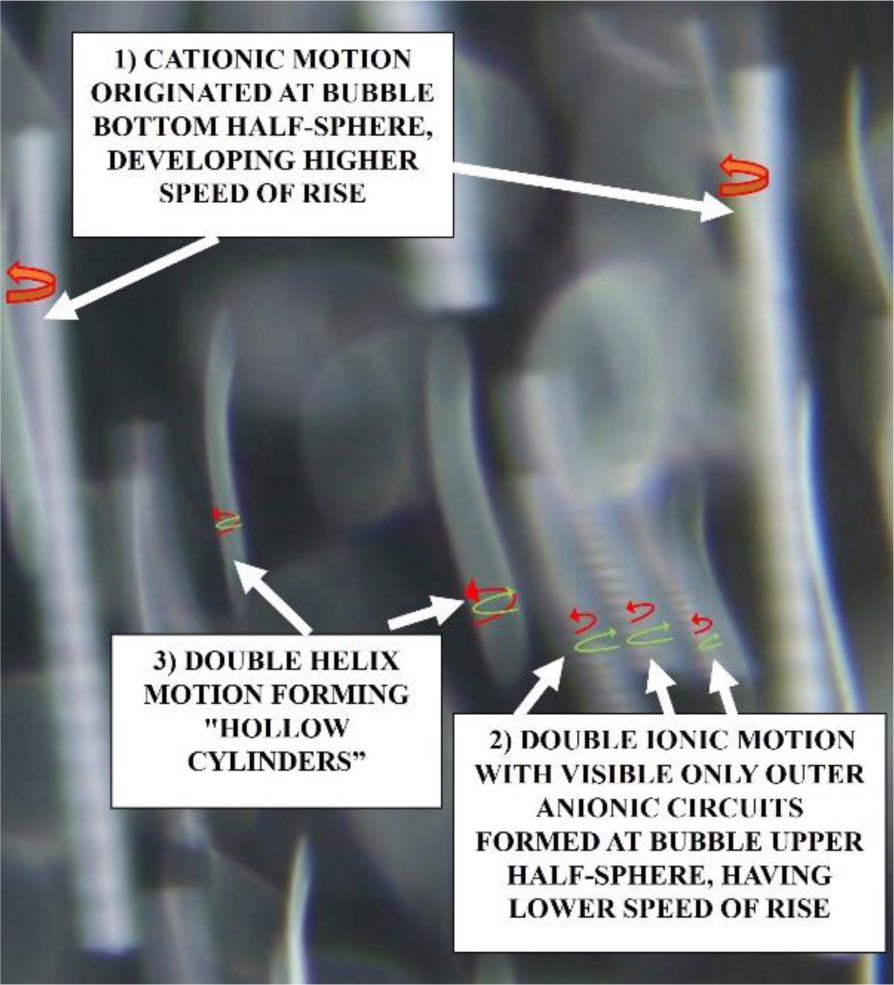
Research dedicated to trace rotational motion of bubbles in saline water revealed that these may generate either single cationic or cationic/anionic motions, including spliced double helix flow. In all cases, the aggregated ionic flows propagate in spiraling as well as rotational manner. However, if bi-ionic or double helix motion is generated, the flow is oppositely directed and has opposite electric charges. Next, the assembled flow is forced to pirouette within the bubble vortex. During that processing the narrowing of spiraling flow takes place and result in increase of revolutions to even millions per second. As a result, a significant friction is induced between revolving ionic hydrates allowing continuous detachment of electrons from covalent atomic shells of electropositive elements. Then, free electrons may be attracted by electronegative elements that are dissolved in seawater. Afterwards, that negatively charged elements may undergo electrical condensation around cationic centers of revolutions. That explain a unique mechanism which operates when negatively charged phosphate compounds and pentagonal blocks found in RNA and DNA as ribose as well as pentagonal rings in nitrogenous bases A and G are being winded. The compensative anionic flow and revolutions may conduct winding of hexagonal blocks found in nitrogenous bases A, G and C, T or U. These assume to gather more positive charge needed to bridge negatively charged sugar molecules in nucleic acids. Thus, the continuity in generation of electronegative compounds and spiral manner of arranging them within the sub-bubble vortices should be regarded as a mechanism responsible for precise, rotational-electric polymerization of elongated macromolecules of RNA/DNA architecture. Reported research refers mainly to physical processes activated by rising bubbles thus should be confronted with other experimental methods used in genetics, microbiology and chemistry.
Citation: Roman Marks. Bubble mediated polymerization of RNA and DNA[J]. AIMS Biophysics, 2022, 9(2): 96-107. doi: 10.3934/biophy.2022009
Research dedicated to trace rotational motion of bubbles in saline water revealed that these may generate either single cationic or cationic/anionic motions, including spliced double helix flow. In all cases, the aggregated ionic flows propagate in spiraling as well as rotational manner. However, if bi-ionic or double helix motion is generated, the flow is oppositely directed and has opposite electric charges. Next, the assembled flow is forced to pirouette within the bubble vortex. During that processing the narrowing of spiraling flow takes place and result in increase of revolutions to even millions per second. As a result, a significant friction is induced between revolving ionic hydrates allowing continuous detachment of electrons from covalent atomic shells of electropositive elements. Then, free electrons may be attracted by electronegative elements that are dissolved in seawater. Afterwards, that negatively charged elements may undergo electrical condensation around cationic centers of revolutions. That explain a unique mechanism which operates when negatively charged phosphate compounds and pentagonal blocks found in RNA and DNA as ribose as well as pentagonal rings in nitrogenous bases A and G are being winded. The compensative anionic flow and revolutions may conduct winding of hexagonal blocks found in nitrogenous bases A, G and C, T or U. These assume to gather more positive charge needed to bridge negatively charged sugar molecules in nucleic acids. Thus, the continuity in generation of electronegative compounds and spiral manner of arranging them within the sub-bubble vortices should be regarded as a mechanism responsible for precise, rotational-electric polymerization of elongated macromolecules of RNA/DNA architecture. Reported research refers mainly to physical processes activated by rising bubbles thus should be confronted with other experimental methods used in genetics, microbiology and chemistry.

| [1] |
Blanchard DC (1963) The electrification of the atmosphere by particles from bubbles in the sea. Prog Oceanogr 1: 73-197. https://doi.org/10.1016/0079-6611(63)90004-1 
|
| [2] |
Monahan EC, Fairall CW, Davidson KL, et al. (1983) Observed inter-relations between 10 m winds, ocean whitecaps and marine aerosols. Q J Roy Meteor Soc 109: 379-392. https://doi.org/10.1002/qj.49710946010
|
| [3] | Blanchard DC, Woodcock AH (1957) Bubble formation and modification in the sea and its meteorological significance. Tellus 9: 145-158. https://doi.org/10.3402/tellusa.v9i2.9094 |
| [4] |
Marks R (1990) Preliminary investigations on the influence of rain on the production, concentration, and vertical distribution of sea salt aerosol. J Geophys Res-Oceans 95: 22299-22304. https://doi.org/10.1029/JC095iC12p22299
|
| [5] | Woolf DK (1997) Bubble rotational features-preliminary investigations exchange, In: Liss, P.S., Duce, R.A., The Sea Surface and Global Change, Cambridge: Cambridge University Press 173-205. |
| [6] | Marks R (2014) Bubble rotational features-preliminary investigations. Oceanography 2: 128. http://doi:10.4172/2332-2632.1000128 |
| [7] | Thorpe SA (2001) Breaking waves and near-surface turbulence, In: Steele, J.H., Thorpe, S.A., Turekian, K.K., Encyclopedia of Ocean Sciences, San Diego: Academic Press 349-351. |
| [8] |
Terray EA, Donelan MA, Agarwal YC, et al. (1996) Estimates of kinetic energy dissipated under breaking waves. J Phys Oceanogr 26: 792-807. https://doi.org/10.1175/1520-0485(1996)026<0792:EOKEDU>2.0.CO;2
|
| [9] | Marks R (2015) Sub-bubble bi-pirouette splicing of cationic and anionic bases as a process of RNA/DNA creation. Oceanography 3: 135. http://doi: 10.4172/2332-2632.1000135 |
| [10] |
Marks R (2019) Water vapor induced airborne rotational features. Meteorol Hydrol Wate 7: 39-47. https://doi.org/10.26491/mhwm/104634
|
| [11] |
Marks R, Górecka E, McCartney K, et al. (2019) Rising bubbles as mechanism for scavenging and aerosolization of diatoms. J Aerosol Sci 128: 79-88. https://doi.org/10.1016/j.jaerosci.2018.12.003
|
| [12] |
Novakov T, Penner JE (1993) Large contribution of organic aerosols to cloud-condensation-nuclei concentrations. Nature 365: 823-826. https://doi.org/10.1038/365823a0
|
| [13] |
Bigg EK, Leck C (2008) The composition of fragments of bubbles bursting at the ocean surface. J Geophys Res-Atmos 113: D11209. https://doi.org/10.1029/2007JD009078
|
| [14] |
Draper DE (2008) RNA folding thermodynamic and molecular descriptions of the role of ions. Biophys J 95: 5489-5495. https://doi.org/10.1529/biophysj.108.131813
|
| [15] |
Kubicki M, Odzimek A, Neska M (2016) Relationship of ground-level aerosol concentration and atmospheric electric field at three observation sites in Arctic, Antarctic and Europe. Atmos Res 178–179: 329-346. https://doi.org/10.1016/j.atmosres.2016.03.029
|
| [16] |
Mainelis G, Willeke K, Baron P, et al. (2001) Electric charges on airborne microorganisms. J Aerosol Sci 32: 1087-1110. https://doi.org/10.1016/S0021-8502(01)00039-8
|
| [17] |
Pawłowski PH (2021) Charged amino acids may promote coronavirus SARS-CoV-2 fusion with the host cell. AIMS Biophys 8: 111-120. https://doi:10.3934/biophy.2021008
|
| [18] |
Lipfert J, Doniach S, Das R, et al. (2014) Understanding of nucleic acid-ion interactions. Annu Rev Biochem 83: 813-841. https://doi.org/10.1146/annurev-biochem-060409-092720
|
| [19] |
Heinemann U, Rose Y (2020) Symmetry in nucleic-acid double helices. Symmetry 12: 737. https://doi.org/10.3390/sym12050737
|
| [20] |
Gregory AC, Zayed AA, Conceição-Neto N, et al. (2019) Marine DNA viral macro- and microdiversity from pole to pole. Cell 177: 1109-1123. https://doi.org/10.1016/j.cell.2019.03.040
|
| [21] |
Suttle CA (2007) Marine viruses—major players in the global ecosystem. Nat Rev Microbiol 5: 801-812. https://doi.org/10.1038/nrmicro1750
|
| [22] |
de Farias ST, dos Santos Junior AP, Rêgo TG, et al. (2017) Origin and evolution of RNA-dependent RNA polymerase. Front Genet 8: 125. https://doi.org/10.3389/fgene.2017.00125
|
| [23] |
dos Santos Junior AP, José MV, de Farias ST (2021) From RNA to DNA: Insights about the transition of informational molecule in the biological systems based on the structural proximity between the polymerases. Biosystems 206: 104442. https://doi.org/10.1016/j.biosystems.2021.104442
|
| [24] |
Mönttinen HAM, Ravantti JJ, Stuart DI, et al. (2014) Automated structural comparisons clarify the phylogeny of the right-hand-shaped polymerases. Mol Biol Evol 31: 2741-2752. https://doi.org/10.1093/molbev/msu219
|






Figures(7)

Roman Marks. Bubble mediated polymerization of RNA and DNA[J]. AIMS Biophysics, 2022, 9(2): 96-107. doi: 10.3934/biophy.2022009







 DownLoad:
DownLoad: