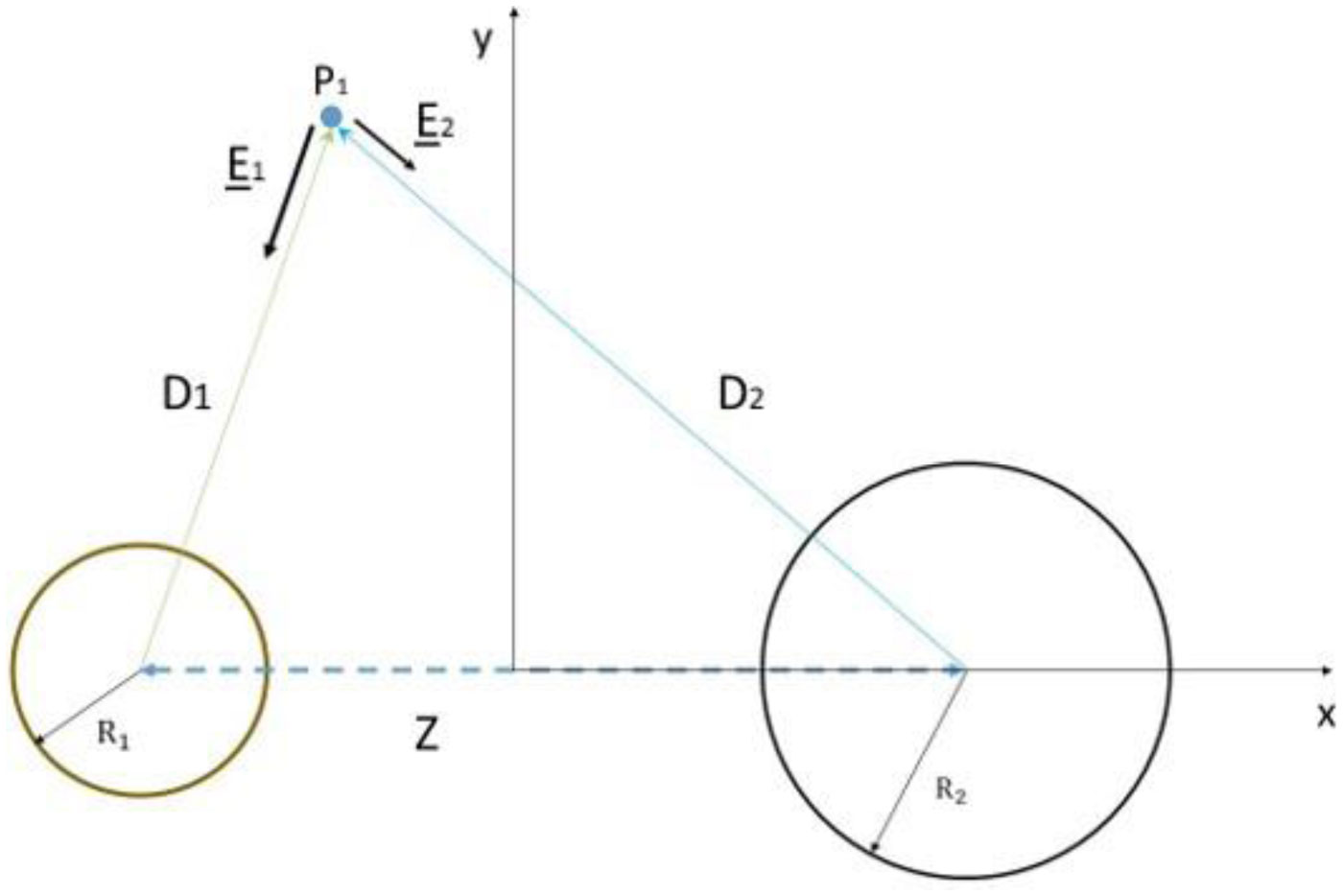
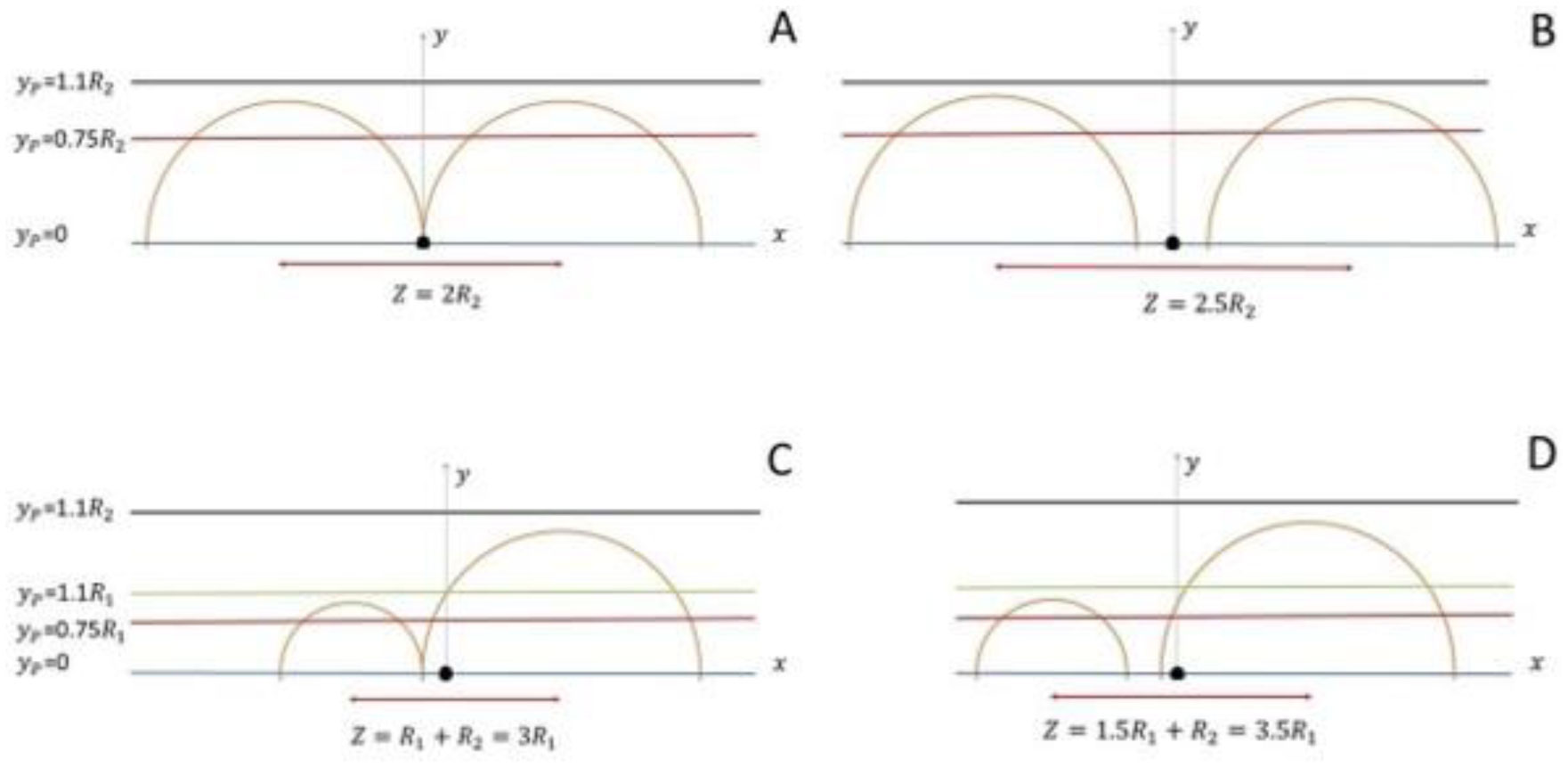
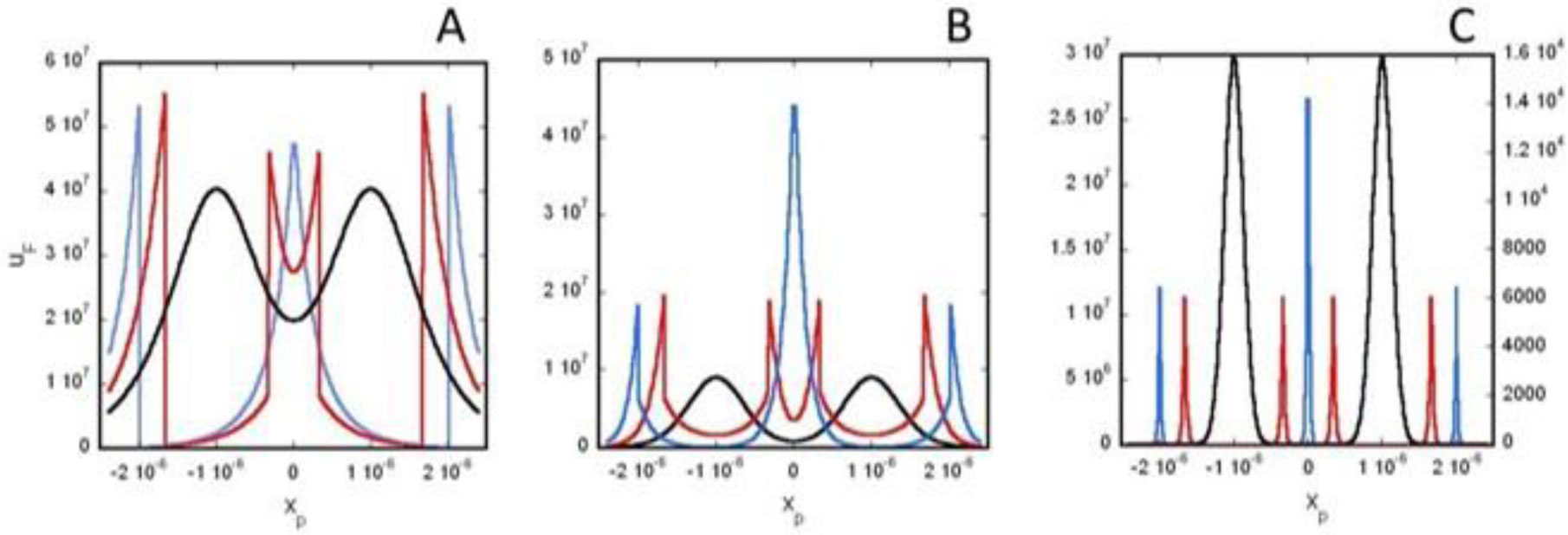
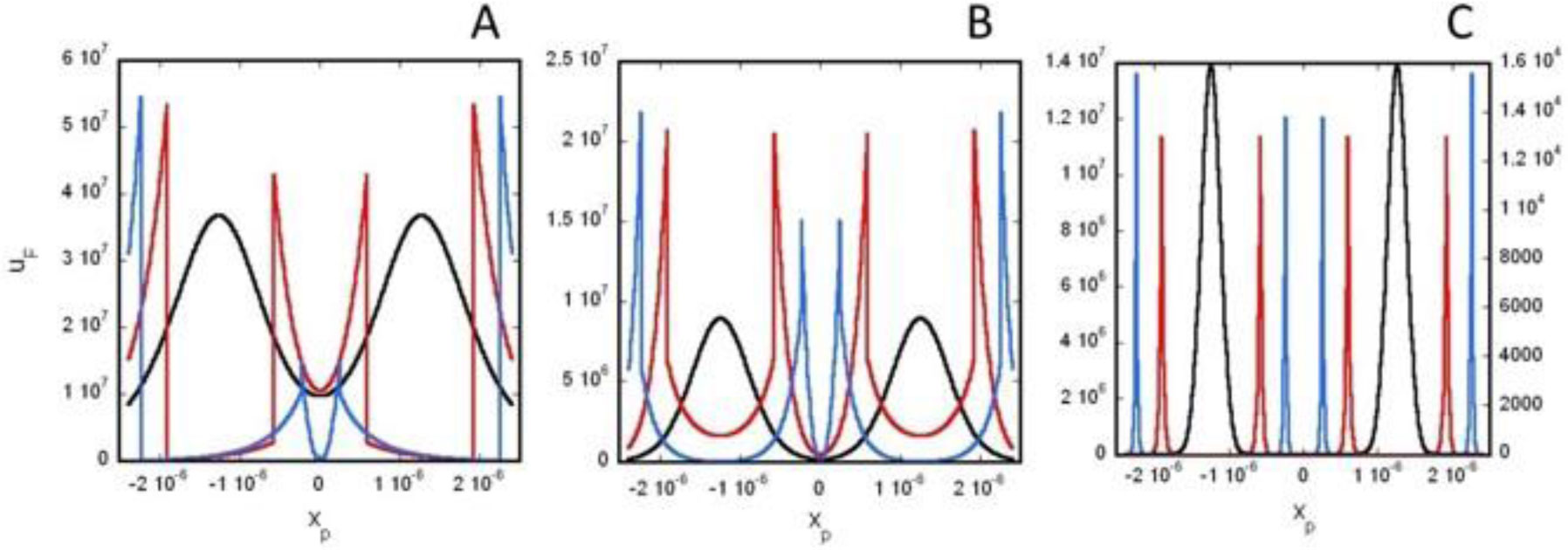
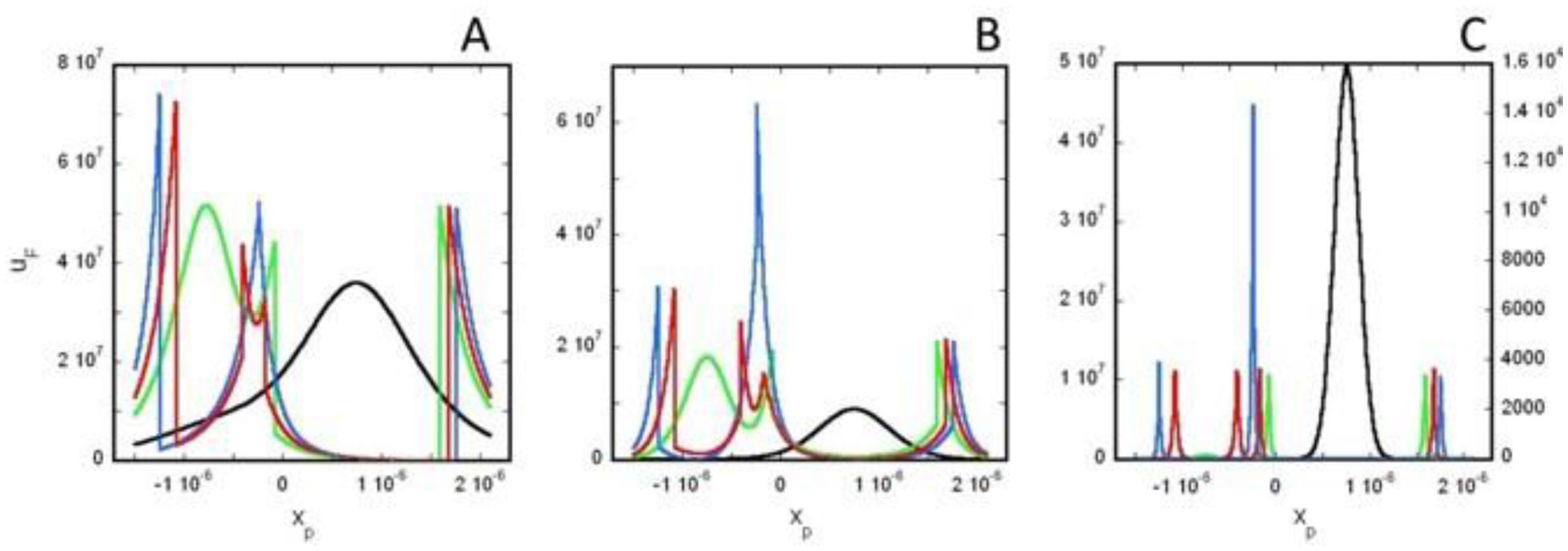
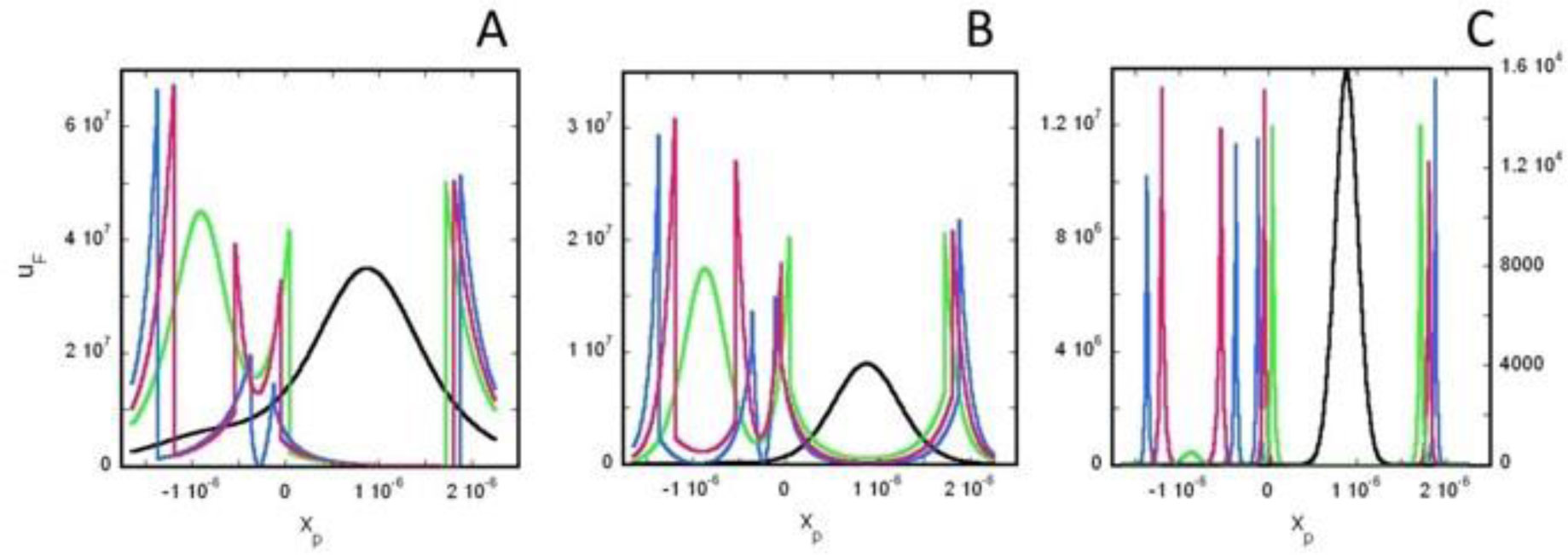
Based on the generalized version of Newton's Shell Theorem the electric field energy density, uF around two separated surface-charged spheres surrounded by electrolyte is calculated. According to the calculations when the surfaces of the charged spheres are farther from each other than four times of the Debye length the field energy density around one of the charged sphere is basically independent from the presence of the other sphere. In this case at low electrolyte ion concentration uF = 0 within the spheres and outside the sphere uF decreases with increasing distance from the surface of the sphere, while at high electrolyte ion concentration uF fast decreases with increasing inner and outer distance from the surface of the sphere. When the charged sheres are close to each other their electric interaction affects the field energy density especially where the surfaces of the spheres are close to each other. Also to model electrophoresis analytical equations are derived for the interaction energy between and the density of electric field energy around a charged flat surface and a charged sphere surrounded by neutral electrolyte.
Citation: István P. Sugár. Density of electric field energy around two surface-charged spheres surrounded by electrolyte I. The spheres are separated from each other[J]. AIMS Biophysics, 2022, 9(2): 86-95. doi: 10.3934/biophy.2022008
Based on the generalized version of Newton's Shell Theorem the electric field energy density, uF around two separated surface-charged spheres surrounded by electrolyte is calculated. According to the calculations when the surfaces of the charged spheres are farther from each other than four times of the Debye length the field energy density around one of the charged sphere is basically independent from the presence of the other sphere. In this case at low electrolyte ion concentration uF = 0 within the spheres and outside the sphere uF decreases with increasing distance from the surface of the sphere, while at high electrolyte ion concentration uF fast decreases with increasing inner and outer distance from the surface of the sphere. When the charged sheres are close to each other their electric interaction affects the field energy density especially where the surfaces of the spheres are close to each other. Also to model electrophoresis analytical equations are derived for the interaction energy between and the density of electric field energy around a charged flat surface and a charged sphere surrounded by neutral electrolyte.

| [1] |
Gabriel JL, Chong PLG (2000) Molecular modeling of archaebacterial bipolar tetraether lipid membranes. Chem Phys Lipids 105: 193-200. https://doi.org/10.1016/S0009-3084(00)00126-2 
|
| [2] |
Chong PLG (2010) Archaebacterial bipolar tetraether lipids: Physico-chemical and membrane properties. Chem Phys Lipids 163: 253-265. https://doi.org/10.1016/j.chemphyslip.2009.12.006
|
| [3] |
Almeida PFF (2009) Thermodynamics of lipid interactions in complex bilayers. BBA-Biomembranes 1788: 72-85. https://doi.org/10.1016/j.bbamem.2008.08.007
|
| [4] |
Sugár IP, Thompson TE, Biltonen RL (1999) Monte Carlo simulation of two-component bilayers: DMPC/DSPC mixtures. Biophys J 76: 2099-2110. https://doi.org/10.1016/S0006-3495(99)77366-2
|
| [5] | Fetter AL, Walecka JD (2003) Theoretical Mechanics of Particles and Continua. New York: Dover Publications 307-310. |
| [6] |
Sugár IP (2020) A generalization of the shell theorem. Electric potential of charged spheres and charged vesicles surrounded by electrolyte. AIMS Biophys 7: 76-89. https://doi.org/10.3934/biophy.2020007
|
| [7] | Newton I (1999) The Principia: Mathematical Principles of Natural Philosophy. Berkeley: University of California Press. |
| [8] |
Sugár IP (2021) Electric energies of a charged sphere surrounded by electrolyte. AIMS Biophys 8: 157-164. https://doi.org/10.3934/biophy.2021012
|
| [9] | Sugár IP (2021) Interaction energy between two separated charged spheres surrounded inside and outside by electrolyte. arXiv:2103.13959 . |
| [10] |
Griffiths DJ (2005) Introduction to electrodynamics. AM J Phys 73: 574. https://doi.org/10.1119/1.4766311
|
| [11] |
Holtzer AM (1954) The collected papers of Peter JW Debye. Interscience, New York-London, 1954. xxi+ 700 pp., $9.50. J Polym Sci 13: 548-548. https://doi.org/10.1002/pol.1954.120137203
|
| [12] |
Levin S, Levin M, Sharp AK, et al. (1983) Theory of electrokinetic behavior of human erythrocytes. Biophys J 42: 127-135. https://doi.org/10.1016/S0006-3495(83)84378-1
|
| [13] | Korohoda W, Wilk A (2008) Cell Electrophoreses–a method for cell separation and research into cell surface properties. Cell Mol Biol Lett 18: 312-326. https://doi.org/10.2478/s11658-008-0004-y |
 biophy-09-02-008-s001.pdf biophy-09-02-008-s001.pdf |
 |
Figures(6)

István P. Sugár. Density of electric field energy around two surface-charged spheres surrounded by electrolyte I. The spheres are separated from each other[J]. AIMS Biophysics, 2022, 9(2): 86-95. doi: 10.3934/biophy.2022008







 DownLoad:
DownLoad: