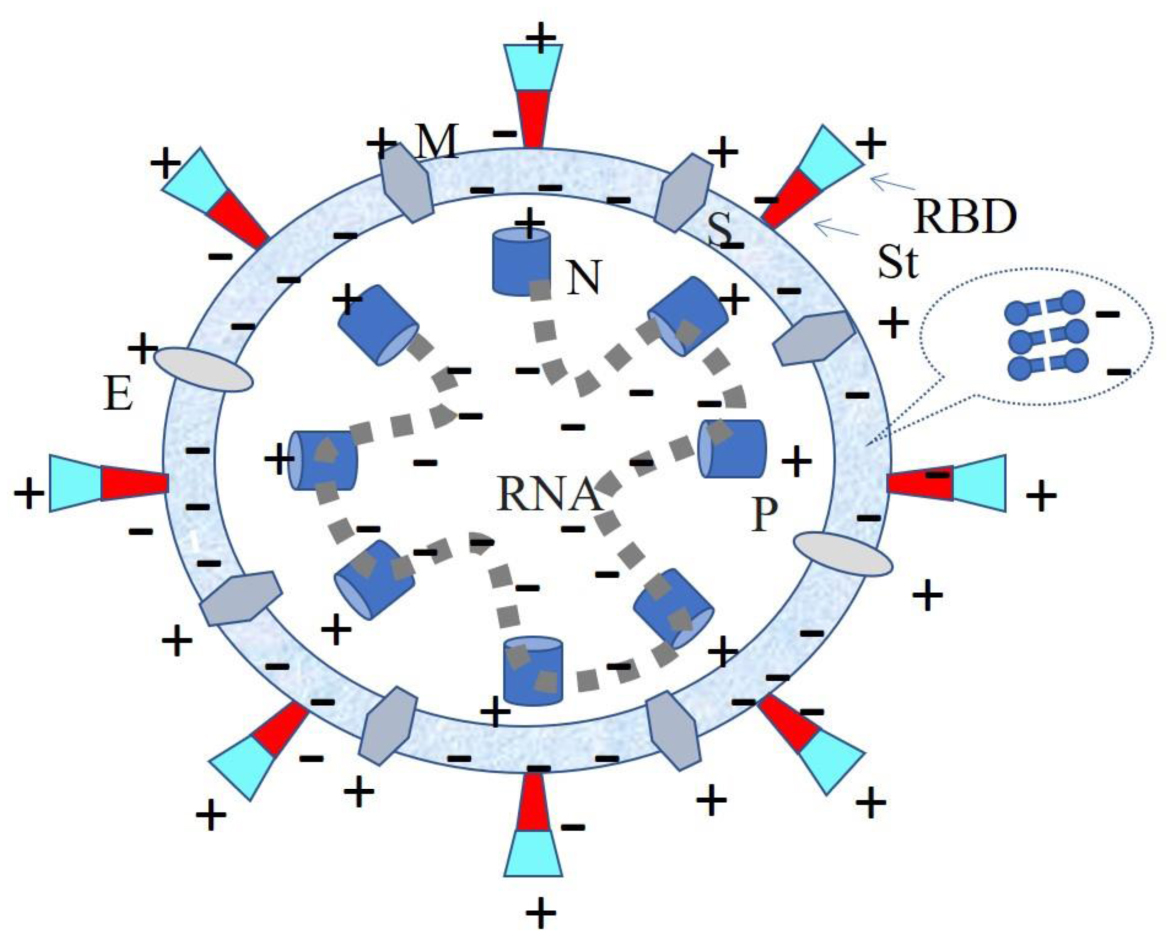
This study analyzed the role of electric charge in human viral infections. Examples of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), dengue, Ebola, influenza A, and respiratory syncytial virus (RSV) are presented. Charge distribution in SARS-CoV-2 and electrostatic interactions of SARS-CoV-2 with its receptor, angiotensin-converting enzyme 2 (ACE2), were evaluated, and the mean time required for respired SARS-CoV-2 virus attachment was evaluated. The virus–cell attachment modality of all of the above viruses was calculated. The impact of electric charge on other viral-related processes, such as replication of virion material, release, and immune response, was also discussed. Special charge conditions in virus treatments were also indicated.
Citation: Piotr H. Pawłowski, Piotr Zielenkiewicz. The role of electric charge in SARS-CoV-2 and other viral infections[J]. AIMS Biophysics, 2024, 11(2): 166-188. doi: 10.3934/biophy.2024011
This study analyzed the role of electric charge in human viral infections. Examples of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), dengue, Ebola, influenza A, and respiratory syncytial virus (RSV) are presented. Charge distribution in SARS-CoV-2 and electrostatic interactions of SARS-CoV-2 with its receptor, angiotensin-converting enzyme 2 (ACE2), were evaluated, and the mean time required for respired SARS-CoV-2 virus attachment was evaluated. The virus–cell attachment modality of all of the above viruses was calculated. The impact of electric charge on other viral-related processes, such as replication of virion material, release, and immune response, was also discussed. Special charge conditions in virus treatments were also indicated.

| [1] | Viral Infections. Physiopedia (2022) . Available from: http:///index.php?title=Viral_Infections&oldid=290753 |
| [2] |
Betts MJ, Russell RB (2003) Amino acid properties and consequences of subsitutions. Bioinformatics for Geneticists . Wiley 289-316. https://doi.org/10.1002/0470867302.ch14 
|
| [3] |
Lipfert J, Doniach S, Das R, et al. (2014) Understanding nucleic acid-ion interactions. Annu Rev Biochem 83: 813-841. https://doi.org/10.1146/annurev-biochem-060409-092720
|
| [4] |
Ma Y, Poole K, Goyette J, et al. (2017) Introducing membrane charge and membrane potential to T cell signaling. Front Immunol 8: 1513. https://doi.org/10.3389/fimmu.2017.01513
|
| [5] |
Zhao X, Ma X, Dupius JH, et al. (2022) Negatively charged phospholipids accelerate the membrane fusion activity of the plant-specific insert domain of an aspartic protease. J Biol Chem 298: 101430. https://doi.org/10.1016/j.jbc.2021.101430
|
| [6] |
Cruz-Chu ER, Malafeev A, Pajarskas T, et al. (2014) Structure and response to flow of the glycocalyx layer. Biophys J 106: 232-242. https://doi.org/10.1016/j.bpj.2013.09.060
|
| [7] | Debye P, Hückel E (1923) The theory of electrolytes. I. Lowering of freezing point and related phenomena (PDF). Physikalische Zeitschrift 24: 185-206. |
| [8] |
Michen B, Graule T (2010) Isoelectric points of viruses. J Appl Microbiol 109: 388-397. https://doi.org/10.1111/j.1365-2672.2010.04663.x
|
| [9] |
Heffron J, Mayer BK (2021) Virus isoelectric point estimation: theories and methods. J Appl Environ Microb 87: e02319-e02320. https://doi.org/10.1128/AEM.02319-20
|
| [10] |
Luisetto M, Tarro G, Edbey K, et al. (2021) Coronavirus COVID-19 surface properties: electrical charges status. Int J Clin Microbiol Biochem Technol 4: 016-027. https://doi.org/10.12688/f1000research.108667.2
|
| [11] |
Cavezzi A, Menicagli R, Troiani E, et al. (2022) COVID-19, cation dysmetabolism, sialic acid, CD147, ACE2, viroporins, hepcidin and ferroptosis: a possible unifying hypothesis. F1000Res 11: 102. https://doi.org/10.12688/f1000research.108667.2
|
| [12] | Northwestern University.“Research exposes new vulnerability for SARS-CoV-2: Electrostatic interactions enhance the spike protein's bond to host cells.”. ScienceDaily (2020) . Available from: https://www.sciencedaily.com/releases/2020/08/200811120227.htm |
| [13] |
Leung WWF, Sun Q (2020) Electrostatic charged nanofiber filter for filtering airborne novel coronavirus (COVID-19) and nano-aerosols. Sep Purif Technol 250: 116886. https://doi.org/10.1016/j.seppur.2020.116886
|
| [14] |
Corrêa Giron C, Laaksonen A, Barroso da Silva FL (2020) On the interactions of the receptor-binding domain of SARS-CoV-1 and SARS-CoV-2 spike proteins with monoclonal antibodies and the receptor ACE2. Virus Res 285: 198021. https://doi.org/10.1016/j.virusres.2020.198021
|
| [15] |
Chavda VP, Bezbaruah R, Deka K, et al. (2022) The delta and omicron variants of SARS-CoV-2: What we know so far. Vaccines 10: 1926. https://doi.org/10.3390/vaccines10111926
|
| [16] | Carabelli AM, Peacock TP, Thorne LG, et al. (2023) SARS-CoV-2 variant biology: immune escape, transmission and fitness. Nat Rev Microbiol 21: 162-177. https://doi.org/10.1038/s41579-022-00841-7 |
| [17] |
Chavda VP, Patel AB, Vaghasiya DD (2022) SARS-CoV-2 variants and vulnerability at the global level. J Med Virol 94: 2986-3005. https://doi.org/10.1002/jmv.27717
|
| [18] |
Chavda VP, Ghali ENHK, Yallapu MM, et al. (2022) Therapeutics to tackle Omicron outbreak. Immunotherapy 14: 833-838. https://doi.org/10.2217/imt-2022-0064
|
| [19] |
Chavda VP, Vuppu S, Mishra T, et al. (2022) Recent review of COVID-19 management: diagnosis, treatment and vaccination. Pharmacol Rep 74: 1120-1148. https://doi.org/10.1007/s43440-022-00425-5
|
| [20] |
Polatoğlu I, Oncu-Oner T, Dalman I, et al. (2023) COVID-19 in early 2023: Structure, replication mechanism, variants of SARS-CoV-2, diagnostic tests, and vaccine & drug development studies. MedComm 4: e228. https://doi.org/10.1002/mco2.228
|
| [21] |
Basu D, Chavda VP, Mehta AA (2022) Therapeutics for COVID-19 and post COVID-19 complications: an update. Curr Res Pharmacol Drug Discov 3: 100086. https://doi.org/10.1016/j.crphar.2022.100086
|
| [22] |
Lancet T (2023) The COVID-19 pandemic in 2023: far from over. Lancet 401: 79. https://doi.org/10.1016/s0140-6736(23)00050-8
|
| [23] |
Arbeitman CR, Rojas P, Ojeda-May P, et al. (2021) The SARS-CoV-2 spike protein is vulnerable to moderate electric fields. Nat Commun 12: 5407. https://doi.org/10.1038/s41467-021-25478-7
|
| [24] |
Božič A, Podgornik R (2024) Changes in total charge on spike protein of SARS-CoV-2 in emerging lineages. Bioinformatics Adv 4: vbae053. https://doi.org/10.1093/bioadv/vbae053
|
| [25] |
Javidpour L, Božič A, Naji A, et al. (2020) Electrostatic interaction between SARS-CoV-2 virus and charged electret fibre. Soft Matter 17: 4296-4303. https://doi.org/10.1039/D1SM00232E
|
| [26] |
Zhang Z, Zhang J, Wang J (2022) Surface charge changes in spike RBD mutations of SARS-CoV-2 and its variant strains alter the virus evasiveness via HSPGs: a review and mechanistic hypothesis. Front Public Health 10: 952916. https://doi.org/10.3389/fpubh.2022.952916
|
| [27] |
Bar-On YM, Flamholz A, Phillips R, et al. (2020) SARS-CoV-2 (COVID-19) by the numbers. elife 9: e57309. https://doi.org/10.7554/eLife.57309
|
| [28] |
Berman HM, Westbrook J, Feng Z, et al. (2000) The protein data bank. Nucleic Acids Res 28: 235-242. https://doi.org/10.1093/nar/28.1.235
|
| [29] |
Sayers EW, Bolton EE, Brister JR, et al. (2022) Database resources of the national center for biotechnology information. Nucleic Acids Res 50: D20-D26. https://doi.org/10.1093/nar/gkab1112
|
| [30] |
Jiao LG, Zan LR, Zhu L, et al. (2019) Accurate computation of screened Coulomb potential integrals in numerical Hartree–Fock programs. Comput Phys Commun 244: 217-227. https://doi.org/10.1016/j.cpc.2019.06.001
|
| [31] | SARS-CoV-2 variants of concern and variants under investigation in England Technical briefing 15. Public Health England (2021) . Available from: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/993879/Variants_of_Concern_VOC_Technical_Briefing_15.pdf |
| [32] | SARS-CoV-2 variants of concern and variants under investigation in England Technical briefing 23. Public Health England, PHE2 (2021) . Available from: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1018547/Technical_Briefing_23_21_09_16.pdf |
| [33] |
Adamczyk Z, Batys P, Barbasz J (2021) SARS-CoV-2 virion physicochemical characteristics pertinent to abiotic substrate attachment. Curr Opin Colloid Interface Sci 55: 101466. https://doi.org/10.1016/j.cocis.2021.101466
|
| [34] |
Berger O, Edholm O, Jähnig F (1997) Molecular dynamics simulations of a fluid bilayer of dipalmitoylphosphatidylcholine at full hydration, constant pressure, and constant temperature. Biophys J 72: 2002-2013. https://doi.org/10.1016/S0006-3495(97)78845-3
|
| [35] |
Krebs F, Scheller C, Grove-Heike K (2021) Isoelectric point determination by imaged CIEF of commercially available SARS-CoV-2 proteins and the hACE2 receptor. Electrophoresis 42: 687-692. https://doi.org/10.1002/elps.202100015
|
| [36] | Pawłowski PH (2021) SARS-CoV-2 variant Omicron (B.1.1.529) is in a rising trend of mutations increasing the positive electric charge in crucial regions of the spike protein S. Acta Biochim Pol 69: 263-264. https://doi.org/10.18388/abp.2020_6072 |
| [37] |
Cotten M, Phan M (2022) Evolution to increased positive charge on the viral spike protein may be part of the adaptation of SARS-CoV-2 to human transmission. iScience 26: 106230. https://doi.org/10.1016/j.isci.2023.106230
|
| [38] |
Lu Y, Zhao T, Lu M, et al. (2021) The analyses of high infectivity mechanism of SARS-CoV-2 and its variants. COVID 1: 666-673. https://doi.org/10.3390/covid1040054
|
| [39] |
Pawłowski PH (2021) Additional positive electric residues in the crucial spike glycoprotein S regions of the new SARS-CoV-2 variants. Infect Drug Resist 14: 5099-5105. https://doi.org/10.2147/IDR.S342068
|
| [40] | Bromage E The risks-know them-avoid them (2020). Available from: https://www.erinbromage.com/post/the-risks-know-them-avoid-them |
| [41] | Islam MA, Ford Versypt AN (2022) Mathematical modeling of impacts of patient differences on COVID-19 lung fibrosis outcomes. bioRxiv [Preprint] . https://doi.org/10.1101/2022.11.06.515367 |
| [42] | Coulomb CA (1785) Premier mémoire sur l'électricité et le magnétisme" [First dissertation on electricity and magnetism]. Histoire de l'Académie Royale des Sciences [History of the Royal Academy of Sciences] (in French) : 569-577. https://www.academie-sciences.fr/pdf/dossiers/Coulomb/Coulomb_pdf/Mem1785_p569.pdf |
| [43] |
Kobayashi Y, Suzuki Y (2012) Compensatory evolution of net-charge in influenza A virus hemagglutinin. PLoS One 7: e40422. https://doi.org/10.1371/journal.pone.0040422
|
| [44] |
Xia S, Liu M, Wang C, et al. (2020) Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res 30: 343-355. https://doi.org/10.1038/s41422-020-0305-x
|
| [45] |
Brunton B, Rogers K, Phillips EK, et al. (2019) TIM-1 serves as a receptor for Ebola virus in vivo, enhancing viremia and pathogenesis. PLoS Negl Trop Dis 13: e0006983. https://doi.org/10.1371/journal.pntd.0006983
|
| [46] | Fahimi H, Allahyari H, Hassan ZM, et al. (2014) Dengue virus type-3 envelope protein domain III; expression and immunogenicity. Iran J Basic Med Sci 11: 836-843. |
| [47] |
Saad-Roy CM, Arinaminpathy N, Wingreen NS, et al. (2020) Implications of localized charge for human influenza A H1N1 hemagglutinin evolution: insights from deep mutational scans. PLoS Comput Biol 16: e1007892. https://doi.org/10.1371/journal.pcbi.1007892
|
| [48] |
Feng Z, Xu L, Xie Z (2022) Receptors for respiratory syncytial virus infection and host factors regulating the life cycle of respiratory syncytial virus. Front Cell Infect Microbiol 12: 858629. https://doi.org/10.3389/fcimb.2022.858629
|
| [49] |
Petrache HI, Tristram-Nagle S, Gawrisch K, et al. (2004) Structure and fluctuations of charged phosphatidylserine bilayers in the absence of salt. Biophys J 86: 1574-1586. https://doi.org/10.1016/S0006-3495(04)74225-3
|
| [50] | Varki A, Schauer R (2009) Sialic Acids. Essentials of Glycobiology . Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press. |
| [51] |
Lupala CS, Lil X, Lei J, et al. (2021) Computational simulations reveal the binding dynamics between human ACE2 and the receptor binding domain of SARS-CoV-2 spike protein. Quant Biol 9: 61-72. https://doi.org/10.15302/J-QB-020-0231
|
| [52] |
Pawłowski PH (2021) Charged amino acids may promote coronavirus SARS-CoV-2 fusion with the host cell. AIMS Biophys 8: 111-121. https://doi.org/10.3934/biophy.2021008
|
| [53] |
Romano M, Ruggiero A, Squeglia F, et al. (2020) A structural view of SARS-CoV-2 RNA replication machinery: RNA synthesis, proofreading and final capping. Cells 9: 1267. https://doi.org/10.3390/cells9051267
|
| [54] |
Silva JRA, Urban J, Araújo E, et al. (2022) Exploring the catalytic mechanism of the RNA cap modification by nsp16-nsp10 complex of SARS-CoV-2 through a QM/MM approach. Int J Mol Sci 23: 300. https://doi.org/10.3390/ijms23010300
|
| [55] |
Kirchdoerfer RN, Ward AB (2019) Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat Commun 10: 2342. https://doi.org/10.1038/s41467-019-10280-3
|
| [56] |
Subissi L, Posthuma CC, Collet A, et al. (2014) One severe acute respiratory syndrome coronavirus protein complex integrates processive RNA polymerase and exonuclease activities. Proc Natl Acad Sc USA 111: E3900-E3909. https://doi.org/10.1073/pnas.1323705111
|
| [57] |
Bianchi M, Borsetti A, Ciccozzi M, et al. (2021) SARS-Cov-2 ORF3a: mutability and function. Int J Biol Macromol 15: 820-826. https://doi.org/10.1016/j.ijbiomac.2020.12.142
|
| [58] |
Scheller C, Krebs F, Minkner R, et al. (2020) Physicochemical properties of SARS-CoV-2 for drug targeting, virus inactivation and attenuation, vaccine formulation and quality control. Electrophoresis 41: 1137-1151. https://doi.org/10.1002/elps.202000121
|
| [59] |
Bohan D, Ert HV, Ruggio N, et al. (2021) Phosphatidylserine receptors enhance SARS-CoV-2 infection. PLoS Pathog 17: e1009743. https://doi.org/10.1371/journal.ppat.1009743
|
| [60] | Alberts B, Johnson A, Lewis J, et al. (2002) Molecular Biology of the Cell. New York: Garland Science. |
| [61] |
Jalloh S, Olejnik J, Berrigan J, et al. (2022) CD169-mediated restrictive SARS-CoV-2 infection of macrophages induces pro-inflammatory responses. PLoS pathogens 18: e1010479. https://doi.org/10.1371/journal.ppat.1010479
|
| [62] |
Watanabe Y, Allen JD, Wrapp D, et al. (2020) Site-specific glycan analysis of the SARS-CoV-2 spike. Science 369: 330-333. https://doi.org/10.1126/science.abb9983
|
| [63] |
von Glasow R, Sander R (2001) Variation of sea salt aerosol pH with relative humidity. Geophys Res Lett 28: 247-250. https://doi.org/10.1029/2000GL012387
|
| [64] |
Field RD, Moelis N, Salzmann J, et al. (2021) Inhaled water and salt suppress respiratory droplet generation and COVID-19 incidence and death on US coastlines. Mol Front J 5.01n02: 17-29. https://doi.org/10.1142/S2529732521400058
|
| [65] |
Duran-Meza AL, Villagrana-Escareño MV, Ruiz-García J, et al. (2021) Controlling the surface charge of simple viruses. PLoS One 16: e0255820. https://doi.org/10.1371/journal.pone.0255820
|
| [66] |
Vega-Acosta JR, Cadena-Nava RD, Gelbart WM, et al. (2014) Electrophoretic mobilities of a viral capsid, its capsid protein, and their relation to viral assembly. J Phys Chem B 118: 1984-1989. https://doi.org/10.1021/jp407379t
|
| [67] |
Bockstahler LE, Kaesberg P (1962) The molecular weight and other biophysical properties of bromegrass mosaic virus. Biophys J 2: 1962. https://doi.org/10.1016/s0006-3495(62)86836-2
|
| [68] |
Johnson MW, Wagner GW, Bancroft JB (1973) A titrimetric and electrophoretic study of cowpea chlorotic mottle virus and its protein. J Gen Virol 19: 263-273. https://doi.org/10.1099/0022-1317-19-2-263
|
| [69] |
van der Schoot P, Bruinsma R (2005) Electrostatics and the assembly of an RNA virus. Phys Rev E 71: 061928. https://doi.org/10.1103/PhysRevE.71.061928
|
| [70] |
Belyi VA, Muthukumar M (2006) Electrostatic origin of the genome packing in viruses. Proc Natl Acad Sci 103: 17174-17178. https://doi.org/10.1073/pnas.0608311103
|
| [71] |
Hagan MF (2009) A theory for viral capsid assembly around electrostatic cores. J Chem Phys 130: 114902. https://doi.org/10.1063/1.3086041
|
| [72] |
Lorenzo-Leal AC, Vimalanathan S, Bach H (2022) Adherence of SARS-CoV-2 delta variant to a surgical mask and N95 respirators. Future Sci OA 8: FSO808. https://doi.org/10.2144/fsoa-2022-0025
|
| [73] |
Javidpour L, Božič A, Najili A, et al. (2021) Electrostatic interaction between SARS-CoV-2 virus and charged electret fibre. Soft Matter 17: 4296-4203. https://doi.org/10.1039/D1SM00232E
|
| [74] |
Ren C, Haghighat F, Feng Z, et al. (2023) Impact of ionizers on prevention of airborne infection in classroom. Build Simul 16: 749-764. https://doi.org/10.1007/s12273-022-0959-z
|
| [75] |
Fantini J, Azzaz F, Chahinian H, et al. (2023) Electrostatic surface potential as a key parameter in virus transmission and evolution: How to manage future virus pandemics in the post-COVID-19 era. Viruses 15: 284. https://doi.org/10.3390/v15020284
|
| [76] |
Wood JP, Magnuson M, Touati A, et al. (2021) Hook Evaluation of electrostatic sprayers and foggers for the application of disinfectants in the era of SARS-CoV-2. PLoS One 16: e0257434. https://doi.org/10.1371/journal.pone.0257434
|
| [77] |
Kalra RS, Kandimalla R (2021) Engaging the spikes: heparan sulfate facilitates SARS-CoV-2 spike protein binding to ACE2 and potentiates viral infection. Sig Transduct Target Ther 6: 39. https://doi.org/10.1038/s41392-021-00470-1
|
| [78] |
Pawłowski P, Szutowicz I, Marszałek P, et al. (1993) Bioelectrorheological model of the cell. 5. Electrodestruction of cellular membrane in alternating electric field. Biophys J 65: 541-549. https://doi.org/10.1016/S0006-3495(93)81056-7
|
| [79] |
Igakura T, Stinchcombe JC, Goon PK, et al. (2003) Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 299: 1713-1716. https://doi.org/10.1126/science.1080115
|








Figures(9) / Tables(2)

Piotr H. Pawłowski, Piotr Zielenkiewicz. The role of electric charge in SARS-CoV-2 and other viral infections[J]. AIMS Biophysics, 2024, 11(2): 166-188. doi: 10.3934/biophy.2024011







 DownLoad:
DownLoad: