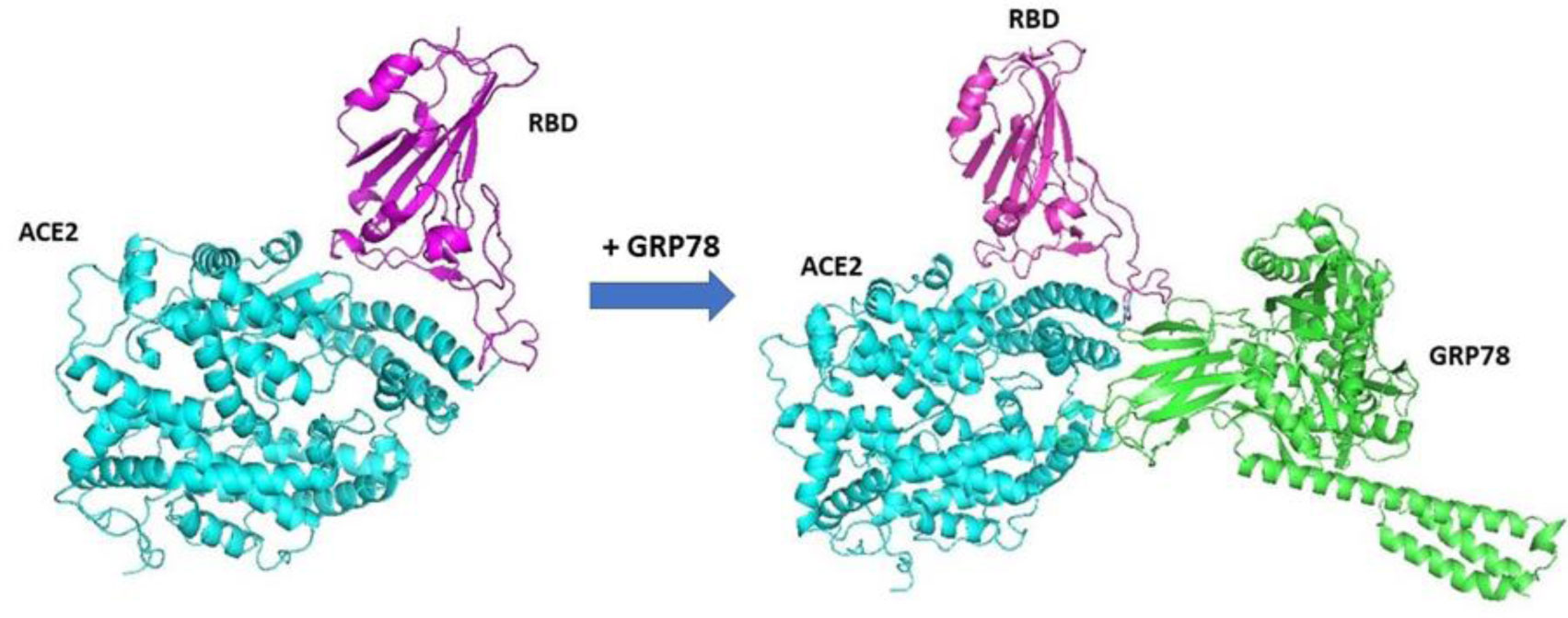
Mucormycosis infection may develop after using steroids treatment to improve the severely of the symptoms in coronavirus patients. The rising in the infection rate of mucormycosis has been noticed in patients after COVID-19 infection. To understand the high morbidity mucormycosis coinfection, the cell surface Glucose Regulated Protein 78 (CS-GRP78) was docked to the virus ACE2-SARS-CoV-2 RBD to create the ACE2-SARS-CoV-2 RBD-GRP78 complex which facilitates the virus entrance into the cell. The spore coat protein homolog 3 (CotH3) of mucormycosis was modeled and docked to the ACE2-SARS-CoV-2 RBD-GRP78 complex. The binding energies of CotH3 with RBD, ACE2, and GRP78 were calculated. The binding results show that GRP78 substrate-binding domain β weakly binds to the spike RBD combined with ACE2 of the spike RBD-ACE2 complex. Its main function is to stabilize the binding between RBD and ACE2, while CotH3 has a strong affinity for the SARS-CoV-2 RBD, but not for ACE2 or GRP78. The CotH3 appeared to have the same affinity to RBD in the SARS-CoV-2 lineages with some preference to the lineage B.1.617.2 (Delta variant). The complex design illustrates that the coat protein of the fungi is more likely linked to the spike protein of the SARS-CoV-2 virus, which would explain the increased mortality mucormycosis coinfections in COVID-19 patients.
Citation: Ibrahim Khater, Aaya Nassar. Looking into mucormycosis coinfections in COVID-19 patients using computational analysis[J]. AIMS Biophysics, 2022, 9(1): 72-85. doi: 10.3934/biophy.2022007
Mucormycosis infection may develop after using steroids treatment to improve the severely of the symptoms in coronavirus patients. The rising in the infection rate of mucormycosis has been noticed in patients after COVID-19 infection. To understand the high morbidity mucormycosis coinfection, the cell surface Glucose Regulated Protein 78 (CS-GRP78) was docked to the virus ACE2-SARS-CoV-2 RBD to create the ACE2-SARS-CoV-2 RBD-GRP78 complex which facilitates the virus entrance into the cell. The spore coat protein homolog 3 (CotH3) of mucormycosis was modeled and docked to the ACE2-SARS-CoV-2 RBD-GRP78 complex. The binding energies of CotH3 with RBD, ACE2, and GRP78 were calculated. The binding results show that GRP78 substrate-binding domain β weakly binds to the spike RBD combined with ACE2 of the spike RBD-ACE2 complex. Its main function is to stabilize the binding between RBD and ACE2, while CotH3 has a strong affinity for the SARS-CoV-2 RBD, but not for ACE2 or GRP78. The CotH3 appeared to have the same affinity to RBD in the SARS-CoV-2 lineages with some preference to the lineage B.1.617.2 (Delta variant). The complex design illustrates that the coat protein of the fungi is more likely linked to the spike protein of the SARS-CoV-2 virus, which would explain the increased mortality mucormycosis coinfections in COVID-19 patients.

| [1] |
Sugar AM (1992) Mucormycosis. Clin Infect Dis 14: S126-S129. https://doi.org/10.1093/clinids/14.Supplement_1.S126 
|
| [2] | Daria S, Asaduzzaman M, Shahriar M, et al. (2021) The massive attack of COVID-19 in India is a big concern for Bangladesh: The key focus should be given on the interconnection between the countries. Int J Health Plan M . https://doi.org/10.1002/hpm.3245 |
| [3] |
Song Y, Qiao J, Giovanni G, et al. (2017) Mucormycosis in renal transplant recipients: review of 174 reported cases. BMC Infect Dis 17: 283. https://doi.org/10.1186/s12879-017-2381-1
|
| [4] |
Bouza E, Muñoz P, Guinea J (2006) Mucormycosis: an emerging disease?. Clin Microbiol Infec 12: 7-23. https://doi.org/10.1111/j.1469-0691.2006.01604.x
|
| [5] |
Farmakiotis D, Kontoyiannis DP (2016) Mucormycoses. Infect Dis Clin N Am 30: 143-163. https://doi.org/10.1016/j.idc.2015.10.011
|
| [6] |
Hamilos G, Samonis G, Kontoyiannis DP (2011) Pulmonary mucormycosis. Semin Respir Crit Care Med 32: 693-702. https://doi.org/10.1055/s-0031-1295717
|
| [7] |
Francis JR, Villanueva P, Bryant P, et al. (2018) Mucormycosis in children: review and recommendations for management. J Pediat Inf Dis Soc 7: 159-164. https://doi.org/10.1093/jpids/pix107
|
| [8] |
Vallabhaneni S, Mody RK (2015) Gastrointestinal mucormycosis in neonates: a review. Curr Fungal Infect Rep 9: 269-274. https://doi.org/10.1007/s12281-015-0239-9
|
| [9] |
Castrejón-Pérez AD, Welsh EC, Miranda I, et al. (2017) Cutaneous mucormycosis. An Bras Dermatol 92: 304-311. https://doi.org/10.1590/abd1806-4841.20176614
|
| [10] |
Spellberg B, Edwards J, Ibrahim A (2005) Novel perspectives on mucormycosis: pathop Mucormycosis: An emerging disease hysiology, presentation, and management. Clin Microbiol Rev 18: 556-569. https://doi.org/10.1128/CMR.18.3.556-569.2005
|
| [11] | Yang W, Shaman J (2021) COVID-19 pandemic dynamics in India and impact of the SARS-CoV-2 Delta (B.1.617.2) variant. MedRxiv . https://doi.org/10.1101/2021.06.21.21259268 |
| [12] | Centers for Disease Control and Preventions, Variants of the Virus. Department of Health & Human Services (2011). Available from: https://www.cdc.gov/coronavirus/2019-ncov/variants/variants.html |
| [13] |
White PL, Dhillon R, Cordey A, et al. (2021) A national strategy to diagnose coronavirus disease 2019–associated invasive fungal disease in the intensive care unit. Clin Infect Dis 73: e1634-e1644. https://doi.org/10.1093/cid/ciaa1298
|
| [14] |
Petrikkos G, Skiada A, Lortholary O, et al. (2012) Epidemiology and clinical manifestations of mucormycosis. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America 54: S23-S34. https://doi.org/10.1093/cid/cir866
|
| [15] |
Drissi C (2021) Black fungus, the darker side of COVID-19. J Neuroradiol 48: 317-318. https://doi.org/10.1016/j.neurad.2021.07.003
|
| [16] |
Bhanuprasad K, Manesh A, Devasagayam E, et al. (2021) Risk factors associated with the mucormycosis epidemic during the COVID-19 pandemic. Int J Infect Dis 111: 267-270. https://doi.org/10.1016/j.ijid.2021.08.037
|
| [17] |
Garg D, Muthu V, Sehgal IS, et al. (2021) Coronavirus disease (Covid-19) associated mucormycosis (CAM): case report and systematic review of literature. Mycopathologia 186: 289-298. https://doi.org/10.1007/s11046-021-00528-2
|
| [18] |
Pal R, Singh B, Bhadada SK, et al. (2021) COVID-19-associated mucormycosis: an updated systematic review of literature. Mycoses 64: 1452-1459. https://doi.org/10.1111/myc.13338
|
| [19] | Kumar M, Sarma DK, Shubham S, et al. (2021) Mucormycosis in COVID-19 pandemic: Risk factors and linkages. Curr Res Microb Sci 2: 100057. https://doi.org/https://doi.org/10.1016/j.crmicr.2021.100057 |
| [20] |
Riad A, Shabaan AA, Issa J, et al. (2021) COVID-19-Associated mucormycosis (CAM): case-series and global analysis of mortality risk factors. J Fungi 7: 837. https://doi.org/10.3390/jof7100837
|
| [21] |
Khatri A, Chang KM, Berlinrut I, et al. (2021) Mucormycosis after Coronavirus disease 2019 infection in a heart transplant recipient–case report and review of literature. J Med Mycol 31: 101125. https://doi.org/10.1016/j.mycmed.2021.101125
|
| [22] |
Singh AK, Singh R, Joshi SR, et al. (2021) Mucormycosis in COVID-19: a systematic review of cases reported worldwide and in India. Diabetes Metab Synd 15: 102146. https://doi.org/10.1016/j.dsx.2021.05.019
|
| [23] |
Devnath P, Dhama K, Tareq AM, et al. (2021) Mucormycosis coinfection in the context of global COVID-19 outbreak: a fatal addition to the pandemic spectrum. Int J Surg (London, England) 92: 106031. https://doi.org/10.1016/j.ijsu.2021.106031
|
| [24] |
Gangneux JP, Bougnoux ME, Dannaoui E, et al. (2020) Invasive fungal diseases during COVID-19: We should be prepared. J Mycol Med 30: 100971. https://doi.org/10.1016/j.mycmed.2020.100971
|
| [25] |
Hoenigl M (2021) Invasive fungal disease complicating coronavirus disease 2019: when it rains, it spores. Clin Infect Dis 73: e1645-e1648. https://doi.org/10.1093/cid/ciaa1342
|
| [26] |
Rabagliati R, Rodríguez N, Núñez C, et al. (2021) COVID-19–associated mold infection in critically Ill patients, Chile. Emerg Infect Dis 27: 1454-1456. https://doi.org/https://doi.org/10.3201/eid2705.204412
|
| [27] |
Ribes JA, Vanover-Sams CL, Baker DJ (2000) Zygomycetes in human disease. Clin Microbiol Rev 13: 236-301. https://doi.org/10.1128/CMR.13.2.236
|
| [28] |
Roden MM, Zaoutis TE, Buchanan WL, et al. (2005) Epidemiology and outcome of zygomycosis: a review of 929 reported cases. Clin Infect Dis 41: 634-653. https://doi.org/10.1086/432579
|
| [29] |
Ibrahim AS, Spellberg B, Walsh TJ, et al. (2012) Pathogenesis of mucormycosis. Clin Infect Dis 54: S16-S22. https://doi.org/10.1093/cid/cir865
|
| [30] |
John TM, Jacob CN, Kontoyiannis DP (2021) When uncontrolled diabetes mellitus and severe COVID-19 converge: the perfect storm for mucormycosis. J Fungi 7: 298. https://doi.org/10.3390/jof7040298
|
| [31] |
Stein DK, Sugar AM (1989) Fungal infections in the immunocompromised host. Diagn Micr Infec Dis 12: 221-228. https://doi.org/https://doi.org/10.1016/0732-8893(89)90140-5
|
| [32] |
Prakash H, Chakrabarti A (2019) Global epidemiology of mucormycosis. J Fungi 5: 26. https://doi.org/10.3390/jof5010026
|
| [33] |
Skiada A, Pavleas I, Drogari-Apiranthitou M (2020) Epidemiology and diagnosis of mucormycosis: an update. J Fungi 6: 265. https://doi.org/10.3390/jof604026
|
| [34] |
Ibrahim AS, Spellberg B, Avanessian V, et al. (2005) Rhizopus oryzae adheres to, is phagocytosed by, and damages endothelial cells in vitro. Infect Immun 73: 778-783. https://doi.org/10.1128/IAI.73.2.778-783.2005
|
| [35] |
Liu M, Spellberg B, Phan QT, et al. (2010) The endothelial cell receptor GRP78 is required for mucormycosis pathogenesis in diabetic mice. J Clin Invest 120: 1914-1924. https://doi.org/10.1172/JCI42164
|
| [36] |
Gumashta J, Gumashta R (2021) COVID19 associated mucormycosis: Is GRP78 a possible link?. J Infect Public Heal 14: 1351-1357. https://doi.org/10.1016/j.jiph.2021.09.004
|
| [37] |
Chandra S, Rawal R (2021) The surge in Covid related mucormycosis. J Infection 83: 381-412. https://doi.org/10.1016/j.jinf.2021.06.008
|
| [38] | Ibrahim AS, Edwards JEJ, Filler SG (2003) Zygomycosis. Clinical Mycology . New York: Oxford University Press 241-251. |
| [39] |
Ibrahim AS (2011) Host cell invasion in mucormycosis: role of iron. Curr Opin Microbiol 14: 406-411. https://doi.org/10.1016/j.mib.2011.07.004
|
| [40] |
Ibrahim AS (2014) Host-iron assimilation: pathogenesis and novel therapies of mucormycosis. Mycoses 57: 13-17. https://doi.org/10.1111/myc.12232
|
| [41] |
Gebremariam T, Liu M, Luo G, et al. (2014) CotH3 mediates fungal invasion of host cells during mucormycosis. J Clin Invest 124: 237-250. https://doi.org/10.1172/JCI71349
|
| [42] |
Alqarihi A, Gebremariam T, Gu Y, et al. (2020) GRP78 and integrins play different roles in host cell invasion during mucormycosis. MBio 11: e01087-20. https://doi.org/10.1128/mBio.01087-20
|
| [43] |
Cuervo NZ, Grandvaux N (2020) ACE2: Evidence of role as entry receptor for SARS-CoV-2 and implications in comorbidities. ELife 9: e61390. https://doi.org/10.7554/eLife.61390
|
| [44] |
Li W, Moore MJ, Vasilieva N, et al. (2003) Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426: 450-454. https://doi.org/10.1038/nature02145
|
| [45] |
Pandiar D, Kumar NS, Anand R, et al. (2021) Does COVID 19 generate a milieu for propagation of mucormycosis?. Med Hypotheses 152: 110613. https://doi.org/10.1016/j.mehy.2021.110613
|
| [46] |
Verma J, Subbarao N (2021) Insilico study on the effect of SARS-CoV-2 RBD hotspot mutants' interaction with ACE2 to understand the binding affinity and stability. Virology 561: 107-116. https://doi.org/10.1016/j.virol.2021.06.009
|
| [47] |
Li J, Lee AS (2006) Stress induction of GRP78/BiP and its role in cancer. Curr Mol Med 6: 45-54. https://doi.org/10.2174/156652406775574523
|
| [48] |
Perlman S, Netland J (2009) Coronaviruses post-SARS: update on replication and pathogenesis. Nat Rev Microbiol 7: 439-450. https://doi.org/10.1038/nrmicro2147
|
| [49] |
Lee AS (2014) Glucose-regulated proteins in cancer: molecular mechanisms and therapeutic potential. Nat Rev Cancer 14: 263-276. https://doi.org/10.1038/nrc3701
|
| [50] |
Ibrahim IM, Abdelmalek DH, Elshahat ME, et al. (2020) COVID-19 spike-host cell receptor GRP78 binding site prediction. J Infection 80: 554-562. https://doi.org/10.1016/j.jinf.2020.02.026
|
| [51] | Ha DP, Van Krieken R, Carlos AJ, et al. (2020) The stress-inducible molecular chaperone GRP78 as potential therapeutic target for coronavirus infection. J Infection 81: 452-482. https://doi.org/10.1016/j.jinf.2020.06.017 |
| [52] |
Elfiky AA, Ibrahim IM, Ismail AM, et al. (2021) A possible role for GRP78 in cross vaccination against COVID-19. J Infection 82: 282-327. https://doi.org/10.1016/j.jinf.2020.09.004
|
| [53] |
Elfiky AA (2020) SARS-CoV-2 spike-heat shock protein A5 (GRP78) recognition may be related to the immersed human coronaviruses. Front Pharmacol 11: 577467. https://doi.org/10.3389/fphar.2020.577467
|
| [54] |
Saghazadeh A, Rezaei N (2020) Towards treatment planning of COVID-19: Rationale and hypothesis for the use of multiple immunosuppressive agents: anti-antibodies, immunoglobulins, and corticosteroids. Int Immunopharmacol 84: 106560. https://doi.org/10.1016/j.intimp.2020.106560
|
| [55] | Carlos AJ, Ha DP, Yeh DW, et al. (2021) GRP78 binds SARS-CoV-2 Spike protein and ACE2 and GRP78 depleting antibody blocks viral entry and infection in vitro. BioRxiv . https://doi.org/10.1101/2021.01.20.427368 |
| [56] |
Weng G, Wang E, Wang Z, et al. (2019) HawkDock: a web server to predict and analyze the protein–protein complex based on computational docking and MM/GBSA. Nucleic Acids Res 47: W322-W330. https://doi.org/10.1093/nar/gkz397
|
| [57] |
Kozakov D, Beglov D, Bohnuud T, et al. (2013) How good is automated protein docking?. Proteins 81: 2159-2166. https://doi.org/10.1002/prot.24403
|
| [58] |
Vajda S, Yueh C, Beglov D, et al. (2017) New additions to the ClusPro server motivated by CAPRI. Proteins 85: 435-444. https://doi.org/https://doi.org/10.1002/prot.25219
|
| [59] |
Kozakov D, Hall DR, Xia B, et al. (2017) The ClusPro web server for protein–protein docking. Nat Protoc 12: 255-278. https://doi.org/10.1038/nprot.2016.169
|
| [60] |
Waterhouse A, Bertoni M, Bienert S, et al. (2018) SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res 46: W296-W303.
|
| [61] |
Bienert S, Waterhouse A, de Beer TAP, et al. (2017) The SWISS-MODEL repository-new features and functionality. Nucleic Acids Res 45: D313-D319.
|
| [62] |
Guex N, Peitsch MC, Schwede T (2009) Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: a historical perspective. Electrophoresis 30: S162-S173. https://doi.org/10.1002/elps.200900140
|
| [63] |
Studer G, Tauriello G, Bienert S, et al. (2021) ProMod3—A versatile homology modelling toolbox. PLoS Comput Biol 17: e1008667. https://doi.org/10.1371/journal.pcbi.1008667
|
| [64] |
Laskowski RA, MacArthur MW, Moss DS, et al. (1993) PROCHECK: a program to check the stereochemical quality of protein structures. J Appl Crystallogr 26: 283-291. https://doi.org/https://doi.org/10.1107/S0021889892009944
|
| [65] |
Laskowski RA, Rullmann JAC, MacArthur MW, et al. (1996) AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J Biomol NMR 8: 477-486. https://doi.org/10.1007/BF00228148
|
| [66] |
Eisenberg D, Lüthy R, Bowie JU (1997) VERIFY3D: assessment of protein models with three-dimensional profiles. Method Enzymol 277: 396-404. https://doi.org/10.1016/s0076-6879(97)77022-8
|
| [67] |
Colovos C, Yeates TO (1993) Verification of protein structures: patterns of nonbonded atomic interactions. Protein Sci 2: 1511-1519. https://doi.org/10.1002/pro.5560020916
|
| [68] | DeLano WL (2002) Pymol: An open-source molecular graphics tool. CCP4 Newsl Protein Crystallogr 40: 82-92. |
| [69] |
Aguiar JA, Tremblay BJM, Mansfield MJ, et al. (2020) Gene expression and in situ protein profiling of candidate SARS-CoV-2 receptors in human airway epithelial cells and lung tissue. Eur Respir J 56: 2001123. https://doi.org/10.1183/13993003.01123-2020
|



Figures(4) / Tables(3)

Ibrahim Khater, Aaya Nassar. Looking into mucormycosis coinfections in COVID-19 patients using computational analysis[J]. AIMS Biophysics, 2022, 9(1): 72-85. doi: 10.3934/biophy.2022007







 DownLoad:
DownLoad: