Citation: Maryam Mazinani, Gholamreza Rezaei Behbehani, Nematollah Gheibi, Alireza Farasat. Study of jack bean urease interaction with luteolin by the extended solvation model and docking simulation[J]. AIMS Biophysics, 2020, 7(4): 429-435. doi: 10.3934/biophy.2020029

| [1] | Kafarski P, Talma M (2018) Recent advances in design of new urease inhibitors: A review. J Adv Res 13: 101-112. |
| [2] | Behbehani GR, Saboury AA, Taleshi E (2018) Determination of partial unfolding enthalpy for lysozyme upon interaction with dodecyltrimethylammonium bromide using an extended solvation model. J Mol Recogn: Interdisciplinary J 21: 132-135. |
| [3] | Shalbafan M, Behbehani GR, Divsalar A (2018) The effect of methotrexate on the structure changes of human serum. Int J 74: 60-67. |
| [4] | Jiang R, Poschet G, Owen R, et al. (2019) Serum concentration of genistein, luteolin and colorectal cancer prognosis. Nutrients 11: 600. |
| [5] | Behbehani GR, Divsalar A, Saboury AA, et al. (2009) A thermodynamic study on the binding of PEG-stearic acid copolymer with lysozyme. J Solution Chem 38: 219. |
| [6] | Trott O, Olson AJ (2010) AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 31: 455-461. |
| [7] | Wallace AC, Laskowski RA, Thornton JM (1995) LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng, Des Sel 8: 127-134. |
| [8] | Zolghadr L, Rezaei Behbehani G (2019) Thermodynamic study of comfarol binding to urease. Orbital: Electron J Chem 11: 151-154. |
| [9] | Mohammadian M, Rezaei Behbehani G, Ghalami-Choobar B, et al. (2019) A thermodynamic study on the binding of polyethyleneglycol 1500 stearic acid with lysozyme. Orbital: Electron J Chem 11: 422-426. |
| [10] | Payeghadr M, Rezaei Behbehani G, Shalbafan M (2017) Lanthanide chlorides decrease the antioxidant property of human serum albumin. Orbital: Electron J Chem 9: 72-75. |
| [11] | Lemeski ET, Behbehani GR, Saboury AA, et al. (2011) Thermodynamic study of the binding of mercury ion to human growth hormone at different temperatures. J Solution Chem 40: 575-586. |
| [12] | Poursoleiman A, Karimi-Jafari MH, Zolmajd-Haghighi Z, et al. (2019) Polymyxins interaction to the human serum albumin: A thermodynamic and computational study. Spectrochim Acta A 217: 155-163. |
| [13] | Jamali Z, Rezaei Behbehani G, Zare K, et al. (2019) Effect of chrysin omega-3 and 6 fatty acid esters on mushroom tyrosinase activity, stability, and structure. J Food Biochem 43: e12728. |
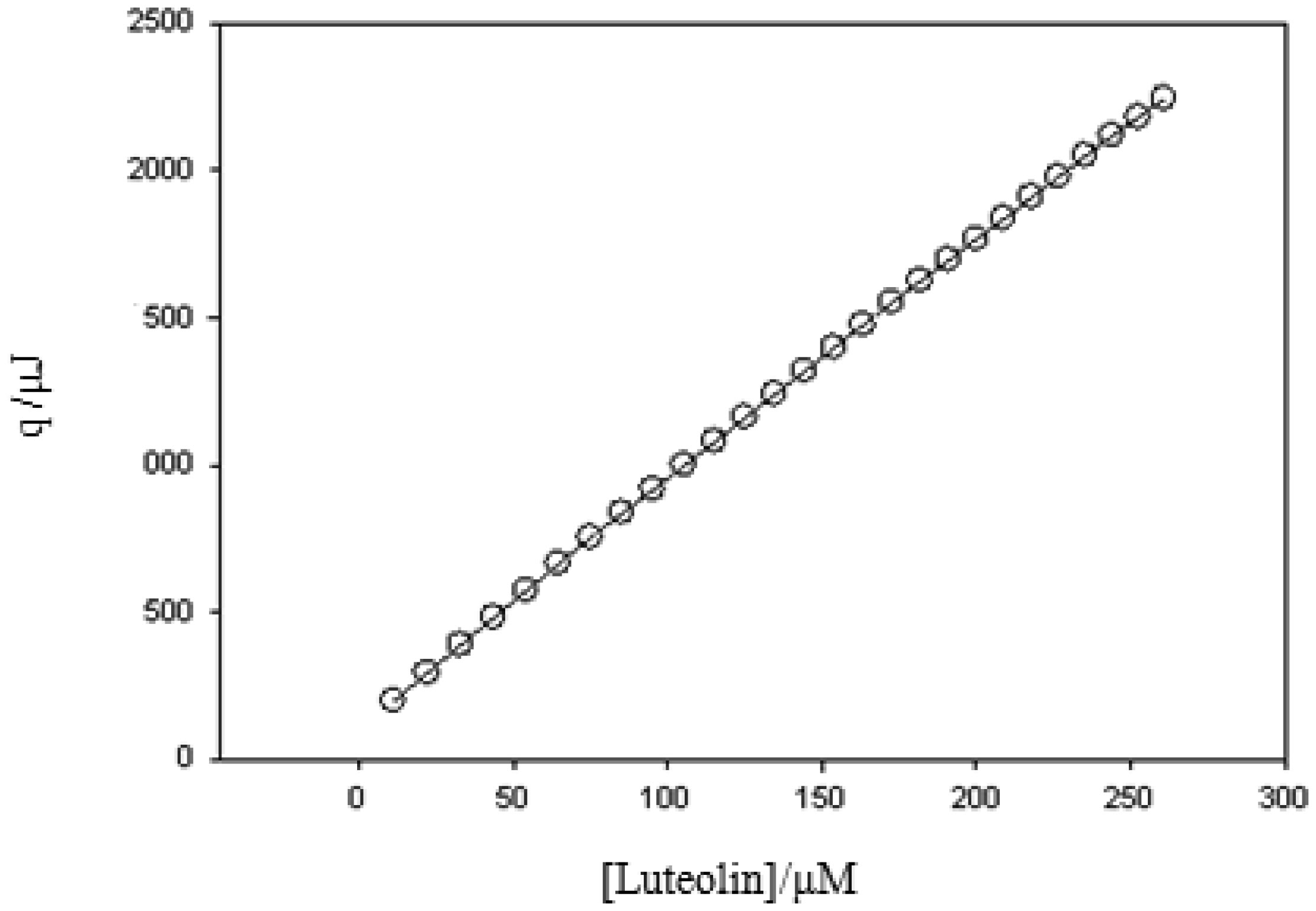
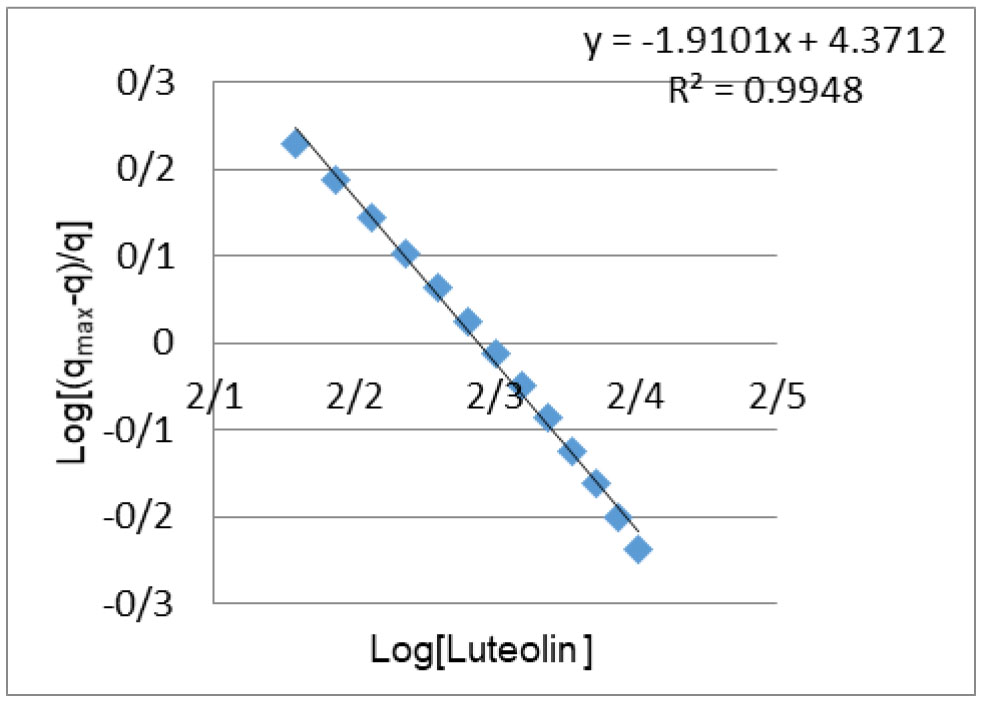
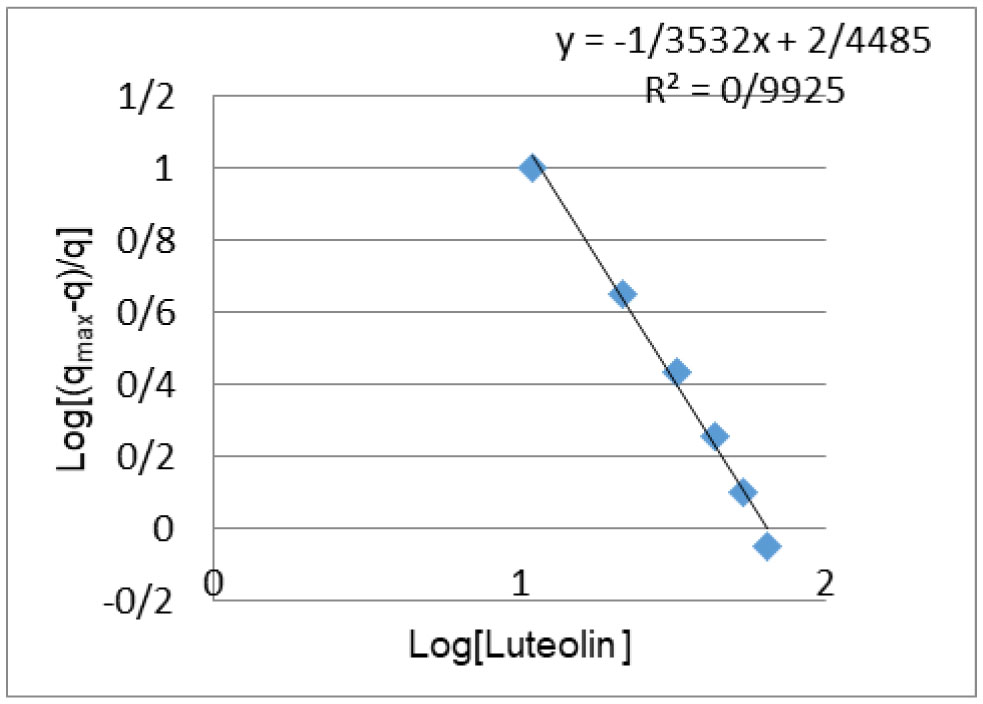

Figures(4) / Tables(2)

Maryam Mazinani, Gholamreza Rezaei Behbehani, Nematollah Gheibi, Alireza Farasat. Study of jack bean urease interaction with luteolin by the extended solvation model and docking simulation[J]. AIMS Biophysics, 2020, 7(4): 429-435. doi: 10.3934/biophy.2020029







 DownLoad:
DownLoad: