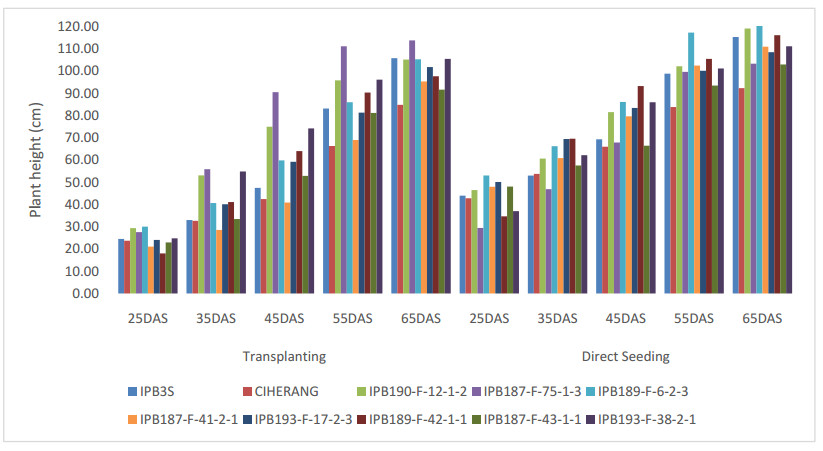
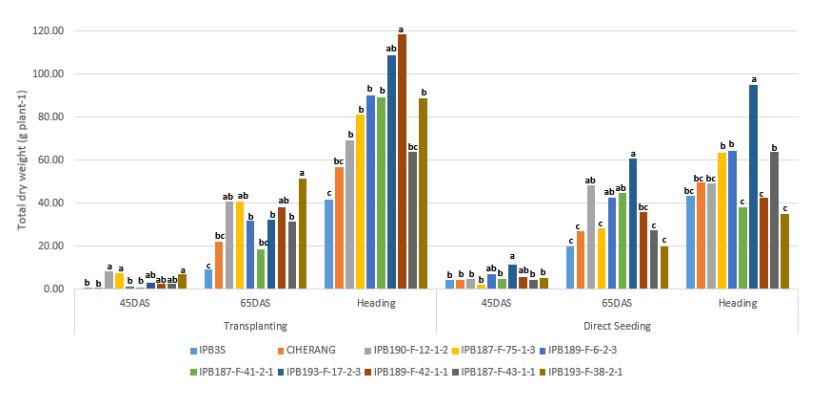
Direct seeding of rice (DSR) may give benefit in using water and labor more efficient and reduce production costs. This study purposes to investigate the character of the seeds, their early vigor traits, the growth and development of rice plants for developing DSR cultivar. The research was conducted in four stages: the measurement of the size of the seed, endosperm, and embryo; the germination test in the laboratory; seedling test using experimental pots; and testing the agronomic performance on transplanting and direct seeding methods in a plastic house. Seed material used eight breeding lines of IPB University and two released varieties. The results of study showed that each genotype had different characteristics of seed, endosperm, and embryo in both weight and area. Seed weight becomes the most dominant in the emergence of superior EV traits, whereas the more seed weight indicates faster radicle emergence and more weight of seedling. DSR method compared to transplanting showed performance such as taller plant, higher leaf area and photosynthesis rate at early growth stage, earlier heading time, and higher plant dry weight since early growth until 65 days old. The seed characters have positively correlation to dry weight of seedlings, number of leaves, leaf area, and canopy dry weight. We also found that higher area of endosperm and embryo significantly correlated to have faster plumule emergence, higher leaf area and plant height. Candidate genotypes for DSR would be further investigated in the field trial agronomically.
Citation: Ahmad Rifqi Fauzi, Ahmad Junaedi, Iskandar Lubis, Munif Ghulamahdi, Hajrial Aswidinnoor, Jun-Ichi Sakagami. Evaluation of rice genotypes on seed attributes and agronomic performance for developing direct-seeded cultivar[J]. AIMS Agriculture and Food, 2022, 7(1): 1-21. doi: 10.3934/agrfood.2022001
Direct seeding of rice (DSR) may give benefit in using water and labor more efficient and reduce production costs. This study purposes to investigate the character of the seeds, their early vigor traits, the growth and development of rice plants for developing DSR cultivar. The research was conducted in four stages: the measurement of the size of the seed, endosperm, and embryo; the germination test in the laboratory; seedling test using experimental pots; and testing the agronomic performance on transplanting and direct seeding methods in a plastic house. Seed material used eight breeding lines of IPB University and two released varieties. The results of study showed that each genotype had different characteristics of seed, endosperm, and embryo in both weight and area. Seed weight becomes the most dominant in the emergence of superior EV traits, whereas the more seed weight indicates faster radicle emergence and more weight of seedling. DSR method compared to transplanting showed performance such as taller plant, higher leaf area and photosynthesis rate at early growth stage, earlier heading time, and higher plant dry weight since early growth until 65 days old. The seed characters have positively correlation to dry weight of seedlings, number of leaves, leaf area, and canopy dry weight. We also found that higher area of endosperm and embryo significantly correlated to have faster plumule emergence, higher leaf area and plant height. Candidate genotypes for DSR would be further investigated in the field trial agronomically.

| [1] |
Rao AN, Johnson DE, Sivaprasad B, et al. (2007) Weed Management in Direct-Seeded Rice. Adv Agron 93: 153-255. https://doi.org/10.1016/S0065-2113(06)93004-1 doi: 10.1016/S0065-2113(06)93004-1

|
| [2] | Pandey S, Velasco L (2005) Trends in crop establishment methods in Asia and research issues, In: Toriyama K, Heong K, Hardy B (Eds.), Rice Is Life: Scientific Perspectives for the 21st Century, International Rice Research Institute, Los Banos, Philippines and Japan International Research Center for Agricultural Sciences, 597. |
| [3] | Quilloy FA, Labaco B, Casal C, et al. (2021) Crop Establishment in Direct-Seeded Rice: Traits, Physiology, and Genetics. https://doi.org/10.1007/978-3-030-66530-2_15 |
| [4] | Pathak H, Tewari AN, Sankhyan S, et al. (2011) Direct-seeded rice: Potential, performance and problems-A review. Curr Adv Agric Sci 3: 77-88. |
| [5] |
Mishra AK, Khanal AR, Pede VO (2017) Is direct seeded rice a boon for economic performance? Empirical evidence from India. Food Policy 73: 10-18. https://doi.org/10.1016/j.foodpol.2017.08.021 doi: 10.1016/j.foodpol.2017.08.021
|
| [6] |
Susilawati HL, Setyanto P, Kartikawati R, et al. (2019) The opportunity of direct seeding to mitigate greenhouse gas emission from paddy rice field. IOP Conf Ser Earth Environ Sci 393: 0-5. https://doi.org/10.1088/1755-1315/393/1/012042 doi: 10.1088/1755-1315/393/1/012042
|
| [7] |
Farooq M, Siddique KHM, Rehman H, et al. (2011) Rice direct seeding: Experiences, challenges and opportunities. Soil Tillage Res 111: 87-98. https://doi.org/10.1016/j.still.2010.10.008 doi: 10.1016/j.still.2010.10.008
|
| [8] | Joshi E, Kumar D, Lal B, et al. (2013) Management of direct seeded rice for enhanced resource - use efficiency. Plant Knowl J 2: 119-134. |
| [9] |
Sandhu N, Yadav S, Singh VK, et al. (2021) Effective crop management and modern breeding strategies to ensure higher crop productivity under direct seeded rice cultivation system: A review. Agronomy 11: 1-25. https://doi.org/10.3390/agronomy11071264 doi: 10.3390/agronomy11071264
|
| [10] |
Dingkuhn M, Schnier H, Datta S, et al. (1990) Diurnal and Developmental Changes in Canopy Gas Exchange in Relation to Growth in Transplanted and Direct-Seeded Flooded Rice. Funct Plant Biol 17: 119. https://doi.org/10.1071/PP9900119 doi: 10.1071/PP9900119
|
| [11] |
Dang X, Thi TGT, Dong G, et al. (2014) Genetic diversity and association mapping of seed vigor in rice (Oryza sativa L.). Planta 239: 1309-1319. https://doi.org/10.1007/s00425-014-2060-z doi: 10.1007/s00425-014-2060-z
|
| [12] |
Ohno H, Banayo NPMC, Bueno CS, et al. (2018) Longer mesocotyl contributes to quick seedling establishment, improved root anchorage, and early vigor of deep-sown rice. F Crop Res 228: 84-92. https://doi.org/10.1016/j.fcr.2018.08.015 doi: 10.1016/j.fcr.2018.08.015
|
| [13] | Balasubramanian V, Hill J (2002) Direct seeding of rice in Asia: emerging issues and strategic research needs for the 21st century, In: Pandey S, Mortimer M, Wade L, et al. (Eds.), Direct seeding: research issues and opportunities. Proceedings of the International Workshop on Direct Seeding in Asian Rice Systems: Strategic Research Issues and Opportunities, International Rice Research Institute, 15-39. |
| [14] | Mackill D, Redona E (1997) Genotype requirements for direct-seeded rice, In: Fukai S, Cooper M, Salisbury J (Eds.), Breeding Strategies for Rainfed Lowland Rice in Drought-prone Environments, Canberra, Australian Centre for International Agricultural Research, 137-143. |
| [15] | Coffman W, Nanda J (1982) Cultivar development for dry-seeded rice, In: Report of a Workshop on: Cropping Systems Research in Asia, Los Banos Laguna, Philliphine, The International Rice Research Institute (IRRI), 149-156. |
| [16] |
Mahender A, Anandan A, Pradhan SK (2015) Early seedling vigour, an imperative trait for direct-seeded rice: An overview on physio-morphological parameters and molecular markers. Planta 241: 1027-1050. https://doi.org/10.1007/s00425-015-2273-9 doi: 10.1007/s00425-015-2273-9
|
| [17] | Diwan J, Channbyregowda M, Shenoy V, et al. (2013) Molecular mapping of early vigour related QTLs in rice. Res J Biol 1: 24-30. |
| [18] | Kumar V, Ladha JK (2011) Direct Seeding of Rice. Recent Developments and Future Research Needs, Elsevier Inc. https://doi.org/10.1016/B978-0-12-387689-8.00001-1 |
| [19] |
Septiningsih EM, Ignacio JCI, Sendon PMD, et al. (2013) QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theor Appl Genet 126: 1357-1366. https://doi.org/10.1007/s00122-013-2057-1 doi: 10.1007/s00122-013-2057-1
|
| [20] |
Huang M, Zhang R, Chen J, et al. (2017) Morphological and physiological traits of seeds and seedlings in two rice cultivars with contrasting early vigor. Plant Prod Sci 20: 95-101. https://doi.org/10.1080/1343943X.2016.1229571 doi: 10.1080/1343943X.2016.1229571
|
| [21] |
Sharma AR (1995) Direct seeding and transplanting for rice production under flood-prone lowland conditions. F Crop Res 44: 129-137. https://doi.org/10.1016/0378-4290(95)00078-X doi: 10.1016/0378-4290(95)00078-X
|
| [22] |
Ohno H, Banayo NPMC, Bueno C, et al. (2018) On-farm assessment of a new early-maturing drought-tolerant rice cultivar for dry direct seeding in rainfed lowlands. F Crop Res 219: 222-228. https://doi.org/10.1016/j.fcr.2018.02.005 doi: 10.1016/j.fcr.2018.02.005
|
| [23] |
Chen C, Jiang Q, Ziska LH, et al. (2015) Seed vigor of contrasting rice cultivars in response to elevated carbon dioxide. F Crop Res 178: 63-68. https://doi.org/10.1016/j.fcr.2015.03.023 doi: 10.1016/j.fcr.2015.03.023
|
| [24] | Pandey M, Seshu D, Akbar M (1992) Genetic variation and association of embryo size to rice seed and seedling vigour. Indian J Genet Plant Breed 52: 310-320. |
| [25] |
Counts RL, Lee PF (1991) Germination and early seedling growth in some northern wild rice (Zizania palustris) populations differing in seed size. Can J Bot 69: 689-696. https://doi.org/10.1139/b91-093 doi: 10.1139/b91-093
|
| [26] |
Namuco OS, Cairns JE, Johnson DE (2009) Investigating early vigour in upland rice (Oryza sativa L.): Part I. Seedling growth and grain yield in competition with weeds. F Crop Res 113: 197-206. https://doi.org/10.1016/j.fcr.2009.05.008 doi: 10.1016/j.fcr.2009.05.008
|
| [27] |
Botwright TL, Condon AG, Rebetzke GJ, et al. (2002) Field evaluation of early vigour for genetic improvement of grain yield in wheat. Aust J Agric Res 53: 1137. https://doi.org/10.1071/AR02007 doi: 10.1071/AR02007
|
| [28] |
Boyd, W.J.R., A.G. Gordon LJL (1971) Seed size, germination resistance and seedling vigor in barley. Can J Plant Sci 51: 93-99. https://doi.org/10.4141/cjps71-021 doi: 10.4141/cjps71-021
|
| [29] |
Fauzi AR, Junaedi A, Lubis I, et al. (2021) Relationship of size and shape rice seed to early seedling vigor traits. IOP Conf Ser Earth Environ Sci 694: 012039. https://doi.org/10.1088/1755-1315/694/1/012039 doi: 10.1088/1755-1315/694/1/012039
|
| [30] |
Shi Z, Chang TG, Chen F, et al. (2020) Morphological and physiological factors contributing to early vigor in the elite rice cultivar 9,311. Sci Rep 10: 1-16. https://doi.org/10.1038/s41598-020-71913-y doi: 10.1038/s41598-020-71913-y
|
| [31] |
Tanabata T, Shibaya T, Hori K, et al. (2012) SmartGrain: High-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol 160: 1871-1880. https://doi.org/10.1104/pp.112.205120 doi: 10.1104/pp.112.205120
|
| [32] | ISTA (2017) International rules for seed testing, The Internastional Seed Testing Association. |
| [33] |
Onwimol D, Chanmprasert W, Changsee P, et al. (2016) Seed vigor classification using analysis of mean radicle emergence time and single counts of radicle emergence in rice (Oryza sativa L.) and mung bean (Vigna radiata (L.) Wilczek). Agric Nat Resour 50: 345-350. https://doi.org/10.1016/j.anres.2016.12.003 doi: 10.1016/j.anres.2016.12.003
|
| [34] |
Morita S, Yonemaru JI, Takanashi JI (2005) Grain growth and endosperm cell size under high night temperatures in rice (Oryza sativa L.). Ann Bot 95: 695-701. https://doi.org/10.1093/aob/mci071 doi: 10.1093/aob/mci071
|
| [35] |
An L, Tao Y, Chen H, et al. (2020) Embryo-Endosperm Interaction and Its Agronomic Relevance to Rice Quality. Front Plant Sci 11: 587641. https://doi.org/10.3389/fpls.2020.587641 doi: 10.3389/fpls.2020.587641
|
| [36] |
Hong SK, Kitano H, Satoh H, et al. (1996) How is embryo size genetically regulated in rice? Development 122: 2051-2058. https://doi.org/10.1242/dev.122.7.2051 doi: 10.1242/dev.122.7.2051
|
| [37] |
Lee G, Piao R, Lee Y, et al. (2019) Identification and Characterization of LARGE EMBRYO, a New Gene Controlling Embryo Size in Rice (Oryza sativa L.). Rice 12: 22. https://doi.org/10.1186/s12284-019-0277-y doi: 10.1186/s12284-019-0277-y
|
| [38] |
Nagasawa N, Hibara KI, Heppard EP, et al. (2013) GIANT EMBRYO encodes CYP78A13, required for proper size balance between embryo and endosperm in rice. Plant J 75: 592-605. https://doi.org/10.1111/tpj.12223 doi: 10.1111/tpj.12223
|
| [39] |
Zhou SR, Yin LL, Xue HW (2013) Functional genomics based understanding of rice endosperm development. Curr Opin Plant Biol 16: 236-246. https://doi.org/10.1016/j.pbi.2013.03.001 doi: 10.1016/j.pbi.2013.03.001
|
| [40] | Pandey, M.P., D.V. Seshu, & M. Akbar (1994) Genetics of embryo size and its relationship with seed and seedling vigour in rice (Oryza sativa L.). Indian J Genet Plant Breed 54: 258-268. |
| [41] |
López-Castañeda C, Richards RA, Farquhar GD, et al. (1996) Seed and seedling characteristics contributing to variation in early vigor among temperate cereals. Crop Sci 36: 1257-1266. https://doi.org/10.2135/cropsci1996.0011183X003600050031x doi: 10.2135/cropsci1996.0011183X003600050031x
|
| [42] |
Cervantes E, Martín JJ, Saadaoui E (2016) Updated Methods for Seed Shape Analysis. Scientifica (Cairo) 2016: 5691825. https://doi.org/10.1155/2016/5691825 doi: 10.1155/2016/5691825
|
| [43] |
Ambika S, Manonmani V, Somasundar G (2014) Review on Effect of Seed Size on Seedling Vigour and Seed Yield. Res J Seed Sci 7: 31-38. https://doi.org/10.3923/rjss.2014.31.38 doi: 10.3923/rjss.2014.31.38
|
| [44] |
Huang R, Jiang L, Zheng J, et al. (2013) Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci 18: 218-226. https://doi.org/10.1016/j.tplants.2012.11.001 doi: 10.1016/j.tplants.2012.11.001
|
| [45] |
Bijanzadeh E, Naderi R, Nosrati K, et al. (2017) Effects of accelerated ageing on germination and biochemistry of eight rice cultivars. J Plant Nutr 40: 156-164. https://doi.org/10.1080/01904167.2016.1201502 doi: 10.1080/01904167.2016.1201502
|
| [46] | Palupi T, Ilyas S, Machmud M, et al. (2016) Effect of Coating Formulation on Viability, Vigor and Storability of Rice Seeds (Oryza sativa L.). J Agron Indones (Indonesian J Agron 40: 21-28. |
| [47] |
Sun J, Wu D, Xu J, et al. (2015) Characterisation of starch during germination and seedling development of a rice mutant with a high content of resistant starch. J Cereal Sci 62: 94-101. https://doi.org/10.1016/j.jcs.2015.01.002 doi: 10.1016/j.jcs.2015.01.002
|
| [48] | Teng, S., B.S. Vergara AAA (1992) Relationship of grain length, width and weight to seedling vigor in rice. Philipp J Crop Sci 17: 17-20. |
| [49] |
Banik NC, Kumar A, Mohapatra BK, et al. (2020) Weed management approaches in direct-seeded rice in eastern Indian ecologies-A critical and updated review. Indian J Weed Sci 52: 1-9. https://doi.org/10.5958/0974-8164.2020.00001.5 doi: 10.5958/0974-8164.2020.00001.5
|
| [50] |
Okami M, Kato Y, Yamagishi J (2013) Grain yield and leaf area growth of direct-seeded rice on flooded and aerobic soils in Japan. Plant Prod Sci 16: 276-279. https://doi.org/10.1626/pps.16.276 doi: 10.1626/pps.16.276
|
| [51] |
Chen S, Cai S guan, Chen X, et al. (2009) Genotypic Differences in Growth and Physiological Responses to Transplanting and Direct Seeding Cultivation in Rice. Rice Sci 16: 143-150. https://doi.org/10.1016/S1672-6308(08)60071-2 doi: 10.1016/S1672-6308(08)60071-2
|
| [52] |
Naklang K, Fukai S, Nathabut K (1996) Growth of rice cultivars by direct seeding and transplanting under upland and lowland conditions. F Crop Res 48: 115-123. https://doi.org/10.1016/S0378-4290(96)01029-5 doi: 10.1016/S0378-4290(96)01029-5
|
Figures(2) / Tables(8)

Ahmad Rifqi Fauzi, Ahmad Junaedi, Iskandar Lubis, Munif Ghulamahdi, Hajrial Aswidinnoor, Jun-Ichi Sakagami. Evaluation of rice genotypes on seed attributes and agronomic performance for developing direct-seeded cultivar[J]. AIMS Agriculture and Food, 2022, 7(1): 1-21. doi: 10.3934/agrfood.2022001







 DownLoad:
DownLoad: