Several studies have demonstrated that depression include disruptions not only for mental human disorders but also their healthy living. Rodent-based behavioral tests and models are widely used to understand the mechanisms by which stress triggers anxiety-related behaviors.
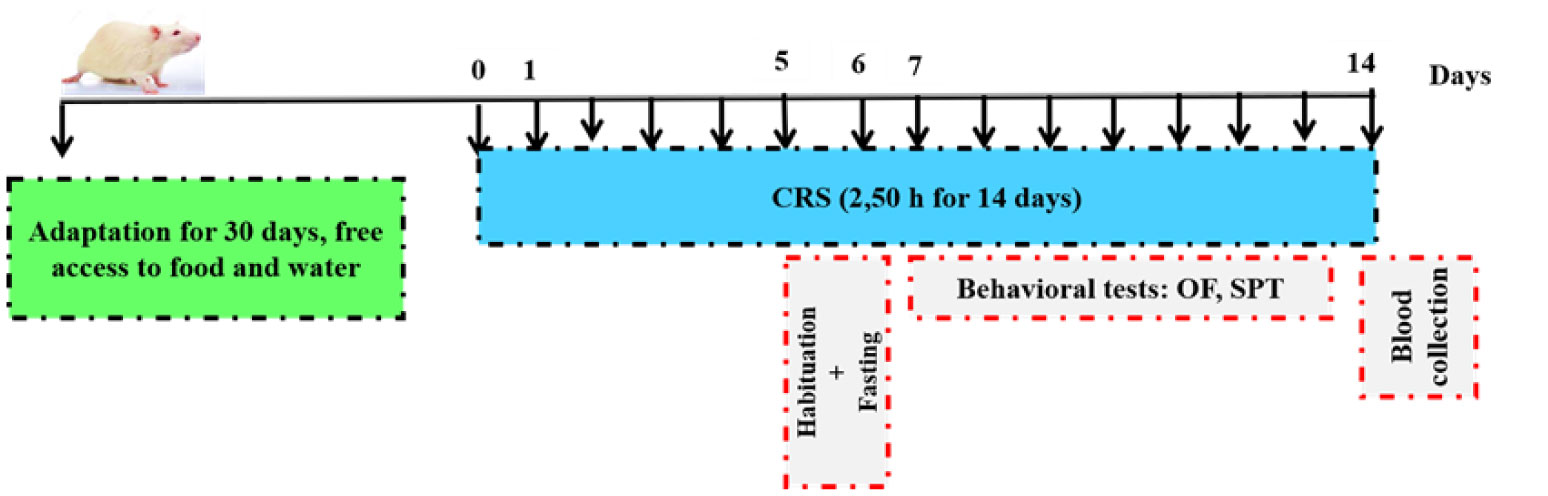
This present study examined the evidence of a chronic restraint stress (CRS) paradigm in male Wistar rats for the progressive nature of depression alongside with related changes in behavior and functions. The body weight was determined, and the behavior tests, including sucrose preference and the open field test were performed. Theses parameters confirme the presence of anxiety-like and depression-like behaviors beside that we will focus on the response of ACTH and testosterone concentrations in rats.
The results obtained during the experiment show that CRS led to decrease the time spent in the field center, a decrease of total distance travelled, in the stressed group compared with the control group. A significant increased of ACTH levels and decreased in testosterone hormone levels in the CRS. According to these results the CRS rodent model has value to validating the development for depression.
Citation: Ahlem Matallah, Rabie Guezi, Abdelmadjid Bairi. Repeated restraint stress induced neurobehavioral and sexual hormone disorders in male rats[J]. AIMS Neuroscience, 2022, 9(2): 264-276. doi: 10.3934/Neuroscience.2022014
Several studies have demonstrated that depression include disruptions not only for mental human disorders but also their healthy living. Rodent-based behavioral tests and models are widely used to understand the mechanisms by which stress triggers anxiety-related behaviors.
This present study examined the evidence of a chronic restraint stress (CRS) paradigm in male Wistar rats for the progressive nature of depression alongside with related changes in behavior and functions. The body weight was determined, and the behavior tests, including sucrose preference and the open field test were performed. Theses parameters confirme the presence of anxiety-like and depression-like behaviors beside that we will focus on the response of ACTH and testosterone concentrations in rats.
The results obtained during the experiment show that CRS led to decrease the time spent in the field center, a decrease of total distance travelled, in the stressed group compared with the control group. A significant increased of ACTH levels and decreased in testosterone hormone levels in the CRS. According to these results the CRS rodent model has value to validating the development for depression.

| [1] |
Smith KE, Pollak SD (2020) Early life stress and development: potential mechanisms for adverse outcomes. J Neurodevel Disord 12: 1-15. https://doi.org/10.1186/s11689-020-09337-y 
|
| [2] |
Shoji H, Miyakawa T (2020) Differential effects of stress exposure via two types of restraint apparatuses on behavior and plasma corticosterone level in inbred male BALB/cAJcl mice. Neuropsychopharmacol Rep 40: 73-84. https://doi.org/10.1002/npr2.12093
|
| [3] | Newsom R (2020) Depression and Sleep. Sleep Foundation 9: 18. |
| [4] | Guy-Evans O (2021) Hypothalamic-Pituitary-Adrenal Axis. Stress Response HPA Axis . |
| [5] | Sapolsky RM, Romero LM, Munck AU (2000) How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21: 55-89. https://doi.org/10.1210/edrv.21.1.0389 |
| [6] |
Heck AL, Handa RJ (2019) Sex differences in the hypothalamic–pituitary–adrenal axis' response to stress: an important role for gonadal hormones. Neuropsychopharmacol 44: 45-58. https://doi.org/10.1038/s41386-018-0167-9
|
| [7] |
Buynitsky T, Mostofsky DI (2009) Restraint stress in biobehavioral research: recent developments. Neurosci Biobehav Rev 33: 1089-98. https://doi.org/10.1016/j.neubiorev.2009.05.004
|
| [8] |
Paré WP, Glavin GB (1986) Restraint stress in biomedical research: a review. Neurosci Biobehav Rev 10: 339-70. https://doi.org/10.1016/0149-7634(86)90017-5
|
| [9] |
Hjollund N, Bonde J, Henriksen T, et al. (2004) Reproductive effects of male psychologic stress. Epidemiology 6: 21-27. https://www.jstor.org/stable/20485836
|
| [10] |
El-Naggar HAEM, El-Safty FENA, El-mehi AE, et al. (2020) Effect of Chronic Stress on The Testis of The Adult Male Albino Rat and The Role of Ginger. Egypt J Hosp Med 81: 2184-2194. https://doi.org/10.21608/ejhm.2020.127968
|
| [11] |
Wingfield J, Sapolsky R (2003) Reproduction and resistance to stress: when and how. J Neuroendocrinol 15: 711-724. https://doi.org/10.1046/j.1365-2826.2003.01033.x
|
| [12] |
Oduwole OO, Peltoketo H, Huhtaniemi IT (2018) Role of follicle-stimulating hormone in spermatogenesis. Front Endocrinol 9: 763. https://doi.org/10.3389/fendo.2018.00763
|
| [13] | Chatterjee A, Rajikin MH, Chatterjee R, et al. (2006) Stress and how it affects reproduction. Biomed Res 17: 1-6. https://doi.org/10.1523/eneuro.0113-20.2020 |
| [14] | Seewoo BJ, Hennessy LA, Feindel KW, et al. (2020) Validation of chronic restraint stress model in rats for the study of depression using longitudinal multimodal MR imaging. bioRxiv . https://doi.org/10.1101/2020.03.20.998195 |
| [15] |
Ulloa JL, Castañeda P, Berríos C, et al. (2010) Comparison of the antidepressant sertraline on differential depression-like behaviors elicited by restraint stress and repeated corticosterone administration. Pharmacol Biochem Behav 97: 213-221. https://doi.org/10.1016/j.pbb.2010.08.001
|
| [16] | Díaz A, León P, Conde C, et al. (2010) Evaluación del efecto de la Administración Aguda de Bromocriptina Sobre el Consumo de Sacarosa en Ratas Sometidas a Aislamiento Social. Colombia: Universidad Pontificia Bolivariana. |
| [17] | Alvarez D (2015) Anhedonia en Perros: Efecto del Estrés Sobre la Preferencia Frente a Sacarosa. Universidad de Chile . Available from: http://repositorio.uchile.cl/bitstream/handle/2250/136219/Anhedonia-en-perros-%20efecto-de-%20estr%C3%A9s-sobre-la-preferencia-frente-a-sacarosa.pdf?sequence=1 |
| [18] | Acero-Castillo MC, Ardila-Figueroa MC, de Oliveira SB (2021) Anhedonic type behavior and anxiety profile of Wistar-UIS rats subjected to chronic social isolation. Front Behav Neurosci 15. https://doi.org/10.3389/fnbeh.2021.663761 |
| [19] |
Klein DF (1974) Endogenomorphic depression: a conceptual and terminological revision. Arch Gen Psychiatry 31: 447-454. https://doi.org/10.1001/archpsyc.1974.01760160005001
|
| [20] | Vidal J (2014) Open field modifications needed to measure, in the mouse, exploration-driven ambulation and fear of open space. Anuario de psicología/ UB J psychol 44: 7-19. http://hdl.handle.net/2445/114794 |
| [21] |
Xu XE, Liu L, Wang Y, et al. (2019) Caspase-1 inhibitor exerts brain-protective effects against sepsisassociated encephalopathy and cognitive impairments in a mouse model of sepsis. Brain Behav Immun 80: 859-870. https://doi.org/10.1016/j.bbi.2019.05.038
|
| [22] |
Denenberg VH (1969) Open-field behavior in the rat: What does it mean?. Ann N Y Acad Sci 159: 852-859. https://doi.org/10.1111/j.1749-6632.1969.tb12983.x
|
| [23] |
Engvall E, Perlman P (1971) Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Immunochem 8: 871-874. https://doi.org/10.1016/0019-2791(71)90454-X
|
| [24] | Servatius RJ, Salameh G, Coyle KM, et al. (2007) Restraint Stress*, Encyclopedia of Stress.. Academic Press. |
| [25] |
Goshen I, Kreisel T, Ben-Menachem-Zidon O, et al. (2008) Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. Mol Psychiatry 13: 717-728. https://doi.org/10.1038/sj.mp.4002055
|
| [26] |
Liu MY, Yin CY, Zhu LJ, et al. (2018) Sucrose preference test for measurement of stressinduced anhedonia in mice. Nat Protoc 13: 1686-1698. https://doi.org/10.1038/s41596-018-0011-z
|
| [27] | Mao Y, Xu Y, Yuan X (2022) Validity of chronic restraint stress for modeling anhedonic-like behavior in rodents: a systematic review and meta-analysis. J Int Medl Res 50: 03000605221075816. https://doi.org/10.1177/03000605221075816 |
| [28] |
Wang Q, Timberlake IIMA, Prall K, et al. (2017) The recent progress in animal models of depression. Prog Neuro-Psychopharmacol Biol Psychiatry 77: 99-109. https://doi.org/10.1016/j.pnpbp.2017.04.008
|
| [29] |
Ampuero E, Luarte A, Santibanez M, et al. (2015) Two Chronic Stress Models Based on Movement Restriction in Rats Respond Selectively to Antidepressant Drugs: Aldolase C As a Potential Biomarker. Int J Neuropsychopharmacol 18: pyv038. https://doi.org/10.1093/ijnp/pyv038
|
| [30] |
Chai C, Jin B, Yan Y, et al. (2021) Anti-depressant effect of Zhi-zi-chi decoction on CUMS mice and elucidation of its signaling pathway. J Ethnopharmacol 266: 113283. https://doi.org/10.1016/j.jep.2020.113283
|
| [31] |
Zhong F, Liu L, Wei JL, et al. (2019) Brain-derived neurotrophic factor precursor in the hippocampus regulates both depressive and anxiety-like behaviors in rats. Front Psychiatry 9: 776. https://doi.org/10.3389/fpsyt.2018.00776
|
| [32] |
Morgan JA (2018) The effects of aerobic exercise on depression-like, anxiety-like, and cognition-like behaviours over the healthy adult lifespan of C57BL/6 mice. Behav Brain Res 337: 193-203. https://doi.org/10.1016/j.bbr.2017.09.022
|
| [33] |
Walsh RN, Cummins RA (1976) The open-field test: a critical review. Psychol Bull 83: 482-504. https://doi.org/10.1037/0033-2909.83.3.482
|
| [34] | Fee C, Prevot T, Misquitta K, et al. (2020) Chronic stress exacerbates acute stress-induced neuronal activation in the anterior cingulate cortex and ventral hippocampus that correlates with behavioral deficits in mice. bioRxiv . https://doi.org/10.1016/j.neuroscience.2020.05.034 |
| [35] |
Orr TE, Mann DR (1990) Effects of restraint stress on plasma LH and testosterone concentrations, Leydig cell LH/hCG receptors, and in vitro testicular steroidogenesis in adult rats. Horm Behav 24: 324-341. https://doi.org/10.1016/0018-506X(90)90013-N
|
| [36] |
SAFYA EE, RADWA M (2018) Effect of Visfatin on Testosterone Hormone Level in Chronic Restraint Male Albino Rats. Med J Cairo Univ 86: 3733-3741. https://doi.org/10.21608/MJCU.2018.61488
|
| [37] |
Bagheri Y, Fathi E, Maghoul A, et al. (2021) Effects of Achillea tenuifolia Lam. hydro-alcoholic extract on anxiety-like behavior and reproductive parameters in rat model of chronic restraint stress. Hum Exp Toxicol 40: 1852-1866. https://doi.org/10.1177/09603271211026723
|
| [38] |
McHenry J, Carrier N, Hull E, et al. (2014) Sex differences in anxiety and depression: role of testosterone. Front Neuroendocrinol 35: 42-57. https://doi.org/10.1016/j.yfrne.2013.09.001
|
| [39] |
Määttänen I, Gluschkoff K, Komulainen K, et al. (2021) Testosterone and specific symptoms of depression: Evidence from NHANES 2011–2016. Compr Psychoneuroendocrinol 6: 100044. https://doi.org/10.1016/j.cpnec.2021.100044
|
| [40] |
Huang Y, Wen J, Chen G (2021) Role and Mechanism of Chronic Restraint Stress in Regulating Energy Metabolism and Reproductive Function Through Hypothalamic Kisspeptin Neurons. J Endoc Soc 5: A551-A552. https://doi.org/10.1210/jendso/bvab048.1122
|
| [41] |
Vaccarino AL, Kastin AJ (2000) Endogenous Opiates. Pept 21: 1975-2034. http://dx.doi.org/10.1016/S0196-9781(00)00345-4
|
| [42] |
Damegh MA (2014) Stress-induced changes in testosterone secretion in male rats: role of oxidative stress and modulation by antioxidants. Open J Anim Sci 4: 70. http://dx.doi.org/10.4236/ojas.2014.42010
|
| [43] |
Retana-Márquez S, Bonilla-Jaime H, Vazquez-Palacios G, et al. (2003) Changes in masculine sexual behavior, corticosterone and testosterone in response to acute and chronic stress in male rats. Horm Behav 44: 327-337. https://doi.org/10.1016/j.yhbeh.2003.04.001
|




Figures(5)

Ahlem Matallah, Rabie Guezi, Abdelmadjid Bairi. Repeated restraint stress induced neurobehavioral and sexual hormone disorders in male rats[J]. AIMS Neuroscience, 2022, 9(2): 264-276. doi: 10.3934/Neuroscience.2022014







 DownLoad:
DownLoad: