Neuropeptide S (NPS), which is a peptide that is involved in the regulation of the stress response, seems to be relevant to the mechanism of action of antidepressants that have anxiolytic properties. However, to date, there have been no reports regarding the effect of long-term treatment with escitalopram or venlafaxine on the NPS system under stress conditions.
This study aimed to investigate the effects of the above-mentioned antidepressants on the NPS system in adult male Wistar rats that were exposed to neonatal maternal separation (MS).
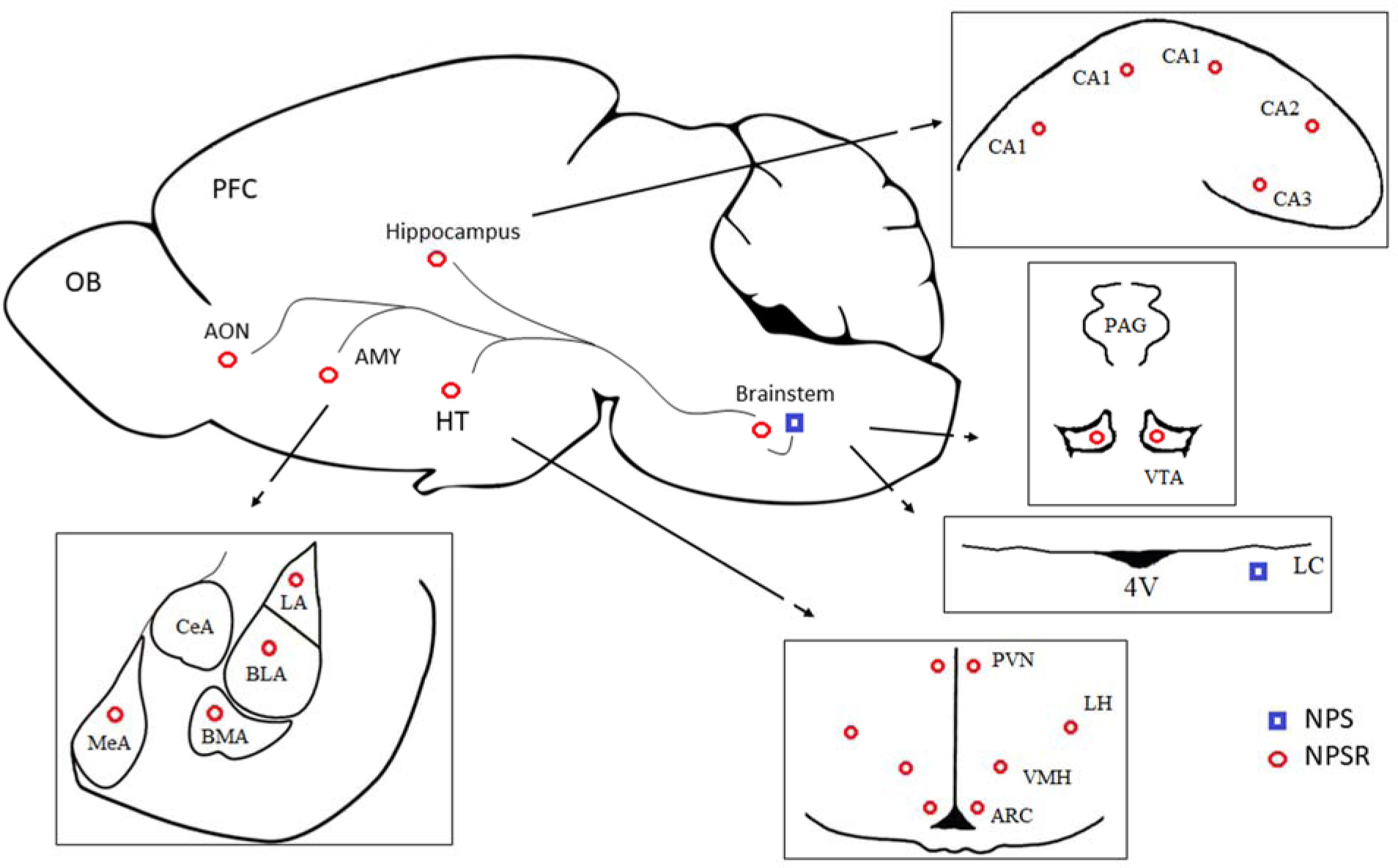
Animals were exposed to MS for 360 min. on postnatal days (PNDs) 2–15. MS causes long-lasting behavioral, endocrine and neurochemical consequences that mimic anxiety- and depression-related features. MS and non-stressed rats were given escitalopram or venlafaxine (10mg/kg) IP from PND 69 to 89. The NPS system was analyzed in the brainstem, hypothalamus, amygdala and anterior olfactory nucleus using quantitative RT-PCR and immunohistochemical methods.
The NPS system was vulnerable to MS in the brainstem and amygdala. In the brainstem, escitalopram down-regulated NPS and NPS mRNA in the MS rats and induced a tendency to reduce the number of NPS-positive cells in the peri-locus coeruleus. In the MS rats, venlafaxine insignificantly decreased the NPSR mRNA levels in the amygdala and a number of NPSR cells in the basolateral amygdala, and increased the NPS mRNA levels in the hypothalamus.
Our data show that the studied antidepressants affect the NPS system differently and preliminarily suggest that the NPS system might partially mediate the pharmacological effects that are induced by these drugs.
Citation: Miłosz Gołyszny, Michał Zieliński, Monika Paul-Samojedny, Artur Pałasz, Ewa Obuchowicz. Chronic treatment with escitalopram and venlafaxine affects the neuropeptide S pathway differently in adult Wistar rats exposed to maternal separation[J]. AIMS Neuroscience, 2022, 9(3): 395-422. doi: 10.3934/Neuroscience.2022022
Neuropeptide S (NPS), which is a peptide that is involved in the regulation of the stress response, seems to be relevant to the mechanism of action of antidepressants that have anxiolytic properties. However, to date, there have been no reports regarding the effect of long-term treatment with escitalopram or venlafaxine on the NPS system under stress conditions.
This study aimed to investigate the effects of the above-mentioned antidepressants on the NPS system in adult male Wistar rats that were exposed to neonatal maternal separation (MS).
Animals were exposed to MS for 360 min. on postnatal days (PNDs) 2–15. MS causes long-lasting behavioral, endocrine and neurochemical consequences that mimic anxiety- and depression-related features. MS and non-stressed rats were given escitalopram or venlafaxine (10mg/kg) IP from PND 69 to 89. The NPS system was analyzed in the brainstem, hypothalamus, amygdala and anterior olfactory nucleus using quantitative RT-PCR and immunohistochemical methods.
The NPS system was vulnerable to MS in the brainstem and amygdala. In the brainstem, escitalopram down-regulated NPS and NPS mRNA in the MS rats and induced a tendency to reduce the number of NPS-positive cells in the peri-locus coeruleus. In the MS rats, venlafaxine insignificantly decreased the NPSR mRNA levels in the amygdala and a number of NPSR cells in the basolateral amygdala, and increased the NPS mRNA levels in the hypothalamus.
Our data show that the studied antidepressants affect the NPS system differently and preliminarily suggest that the NPS system might partially mediate the pharmacological effects that are induced by these drugs.

| [1] |
Kormos V, Gaszner B (2013) Role of neuropeptides in anxiety, stress, and depression: from animals to humans. Neuropeptides 47: 401-419. https://doi.org/10.1016/j.npep.2013.10.014 
|
| [2] |
Yamazoe M, Shiosaka S, Emson PC, et al. (1985) Distribution of neuropeptide Y in the lower brainstem: an immunohistochemical analysis. Brain Res 335: 109-120. https://doi.org/10.1016/0006-8993(85)90281-1
|
| [3] |
Douglas FL, Palkovits M, Brownstein MJ (1982) Regional distribution of substance P-like immunoreactivity in the lower brainstem of the rat. Brain Res 245: 376-378. https://doi.org/10.1016/0006-8993(82)90821-6
|
| [4] |
Spirovski D, Li Q, Pilowsky PM (2012) Brainstem galanin-synthesizing neurons are differentially activated by chemoreceptor stimuli and represent a subpopulation of respiratory neurons. J Comp Neurol 520: 154-173. https://doi.org/10.1002/cne.22723
|
| [5] |
Liu C, Lovenberg TW (2008) Relaxin-3, INSL5, and their receptors. Orphan G Protein-Coupled Receptors and Novel Neuropeptides, 46. Berlin, Heidelberg: Springer 213-237. https://doi.org/10.1007/400_2007_055
|
| [6] |
Palasz A, Rojczyk E, Golyszny M, et al. (2016) Long-term treatment with haloperidol affects neuropeptide S and NPSR mRNA levels in the rat brain. Acta Neuropsychiatr 28: 110-116. https://doi.org/10.1017/neu.2015.56
|
| [7] |
Pałasz A, Żarczyński P, Bogus K, et al. (2021) Modulatory effect of olanzapine on SMIM20/phoenixin, NPQ/spexin and NUCB2/nesfatin-1 gene expressions in the rat brainstem. Pharmacol Rep 73: 1188-1194. https://doi.org/10.1007/s43440-021-00267-7
|
| [8] |
Kumsta R, Chen FS, Pape HC, et al. (2013) Neuropeptide S receptor gene is associated with cortisol responses to social stress in humans. Biol Psychol 93: 304-307. https://doi.org/10.1016/j.biopsycho.2013.02.018
|
| [9] |
Smith KL, Patterson M, Dhillo WS, et al. (2006) Neuropeptide S stimulates the hypothalamo-pituitary-adrenal axis and inhibits food intake. Endocrinology 147: 3510-3518. https://doi.org/10.1210/en.2005-1280
|
| [10] |
Pape HC, Jüngling K, Seidenbecher T, et al. (2010) Neuropeptide S: a transmitter system in the brain regulating fear and anxiety. Neuropharmacology 58: 29-34. https://doi.org/10.1016/j.neuropharm.2009.06.001
|
| [11] |
Xu YL, Gall CM, Jackson VR, et al. (2007) Distribution of neuropeptide S receptor mRNA and neurochemical characteristics of neuropeptide S-expressing neurons in the rat brain. J Comp Neurol 500: 84-102. https://doi.org/10.1002/cne.21159
|
| [12] |
Leonard SK, Ring RH (2011) Immunohistochemical localization of the neuropeptide S receptor in the rat central nervous system. Neuroscience 172: 153-163. https://doi.org/10.1016/j.neuroscience.2010.10.020
|
| [13] |
Nauta WJ (1958) Hippocampal projections and related neural pathways to the midbrainin the cat. Brain 81: 319-340. https://link.springer.com/chapter/10.1007/978-1-4684-7920-1_9
|
| [14] | Nauta WJ, Domesick VB (1982) Neural associations of the limbic system. Dordrecht: Springer 175-206. https://doi.org/10.1007/978-94-011-6302-6_10 |
| [15] |
Soudryl Y, Lemogne C, Malinvaud D, et al. (2011) Olfactory system and emotion: common substrates. Eur Ann Otorhinolaryngol Head Neck Dis 128: 18-23. https://doi.org/10.1016/j.anorl.2010.09.007
|
| [16] |
Xu YL, Reinscheid RK, Huitron-Resendiz S, et al. (2004) Neuropeptide S: a neuropeptide promoting arousal and anxiolytic-like effects. Neuron 43: 487-497. https://doi.org/10.1016/j.neuron.2004.08.005
|
| [17] |
Clark SD, Duangdao DM, Schulz S, et al. (2011) Anatomical characterization of the neuropeptide S system in the mouse brain by in situ hybridization and immunohistochemistry. J Comp Neurol 519: 1867-1893. https://doi.org/10.1002/cne.22606
|
| [18] |
Si W, Aluisio L, Okamura N, et al. (2010) Neuropeptide S stimulates dopaminergic neurotransmission in the medial prefrontal cortex. J Neurochem 115: 475-482. https://doi.org/10.1111/j.1471-4159.2010.06947.x
|
| [19] |
Mochizuki T, Kim J, Sasaki K (2010) Microinjection of neuropeptide S into the rat ventral tegmental area induces hyperactivity and increases extracellular levels of dopamine metabolites in the nucleus accumbens shell. Peptides 31: 926-931. https://doi.org/10.1016/j.peptides.2010.02.006
|
| [20] |
Raiteri L, Luccini E, Romei C, et al. (2009) Neuropeptide S selectively inhibits the release of 5-HT and noradrenaline from mouse frontal cortex nerve endings. Br J Pharmacol 157: 474-481. https://doi.org/10.1111/j.1476-5381.2009.00163.x
|
| [21] |
Pałasz A, Rojczyk E (2015) Neuroleptics affect neuropeptide S and NPSR mRNA levels in the rat brain. J Mol Neurosci 57: 352-357. https://doi.org/10.1007/s12031-015-0625-3
|
| [22] |
Gołyszny M, Obuchowicz E (2019) Are neuropeptides relevant for the mechanism of action of SSRIs?. Neuropeptides 75: 1-17. https://doi.org/10.1016/j.npep.2019.02.002
|
| [23] | Laux G (2021) Serotonin Reuptake Inhibitors: Citalopram, Escitalopram, Fluoxetine, Fluvoxamine, Paroxetine, and Sertraline. NeuroPsychopharmacotherapy . Cham.: Springer 1-13. https://doi.org/10.1007/978-3-319-56015-1_413-1 |
| [24] |
Feng RF, Ma R, Wang P, et al. (2022) Efficacy of escitalopram for poststroke depression: a systematic review and meta-analysis. Sci Rep 12: 1-10. https://doi.org/10.1038/s41598-022-05560-w
|
| [25] |
Zhong H, Haddjeri N, Sánchez C (2012) Escitalopram, an antidepressant with an allosteric effect at the serotonin transporter—a review of current understanding of its mechanism of action. Psychopharmacology 219: 1-13. https://doi.org/10.1007/s00213-011-2463-5
|
| [26] |
Pałasz A, Suszka-Świtek A, Filipczyk Ł, et al. (2016) Escitalopram affects spexin expression in the rat hypothalamus, hippocampus and striatum. Pharmacol Rep 68: 1326-1331. https://doi.org/10.1016/j.pharep.2016.09.002
|
| [27] |
Pałasz A, Piwowarczyk-Nowak A, Suszka-Świtek A, et al. (2020) Escitalopram as a modulator of proopiomelanocortin, kisspeptin, Kiss1R and MCHR1 gene expressions in the male rat brain. Mol Biol Rep 47: 8273-8278. https://doi.org/10.1007/s11033-020-05806-8
|
| [28] | Gołyszny M, Zieliński M, Paul-Samojedny M, et al. (2022) Escitalopram alters the hypothalamic OX system but does not affect its up-regulation induced by early-life stress in adult rats. Neurosci Res . Ahead of print. https//doi.org/10.1016/j.neures.2022.02.005 |
| [29] |
Stahl SM, Morrissette DA, Faedda G, et al. (2017) Guidelines for the recognition and management of mixed depression. CNS Spectrums 22: 203-219. https://doi.org/10.1017/S1092852917000165
|
| [30] | Roseboom PH, Kalin NH (2000) Neuropharmacology of venlafaxine. DepressAnxiety 12: 20-29. https://doi.org/10.1002/1520-6394(2000)12:1+<20::AID-DA3>3.0.CO;2-M |
| [31] |
Stout SC, Owens MJ, Nemeroff CB (2002) Regulation of corticotropin-releasing factor neuronal systems and hypothalamic-pituitary-adrenal axis activity by stress and chronic antidepressant treatment. J Pharmacol Exp Ther 300: 1085-1092. https://doi.org/10.1124/jpet.300.3.1085
|
| [32] |
Petschner P, Juhasz G, Tamasi V, et al. (2016) Chronic venlafaxine treatment fails to alter the levels of galanin system transcripts in normal rats. Neuropeptides 57: 65-70. https://doi.org/10.1016/j.npep.2016.01.010
|
| [33] |
Poretti MB, Sawant RS, Rask-Andersen M, et al. (2016) Reduced vasopressin receptors activation mediates the anti-depressant effects of fluoxetine and venlafaxine in bulbectomy model of depression. Psychopharmacology 233: 1077-1086. https://doi.org/10.1007/s00213-015-4187-4
|
| [34] |
LeMoult J, Humphreys KL, Tracy A, et al. (2020) Meta-analysis: exposure to early life stress and risk for depression in childhood and adolescence. J Am Acad Child Adolesc Psychiatry 59: 842-855. https://doi.org/10.1016/j.jaac.2019.10.011
|
| [35] |
Pervanidou P, Makris G, Chrousos G, et al. (2020) Early life stress and pediatric posttraumatic stress disorder. Brain Sci 10: 169. https://doi.org/10.3390/brainsci10030169
|
| [36] |
Arborelius L, Hawks BW, Owens MJ, et al. (2004) Increased responsiveness of presumed 5-HT cells to citalopram in adult rats subjected to prolonged maternal separation relative to brief separation. Psychopharmacology 176: 248-255. https://doi.org/10.1007/s00213-004-1883-x
|
| [37] |
Lippmann M, Bress A, Nemeroff CB, et al. (2007) Long-term behavioural and molecular alterations associated with maternal separation in rats. Eur J Neurosci 25: 3091-3098. https://doi.org/10.1111/j.1460-9568.2007.05522.x
|
| [38] | Roque S, Mesquita AR, Palha JA, et al. (2014) The behavioral and immunological impact of maternal separation: a matter of timing. Front BehavNeurosci 8: 192. https://doi.org/10.3389/fnbeh.2014.00192 |
| [39] | Bonapersona V, Joels M, Sarabdjitsingh RA (2018) Effects of early life stress on biochemical indicators of the dopaminergic system: A 3 level meta-analysis of rodent studies. NeurosciBiobehav Rev 95: 1-16. https://doi.org/10.1016/j.neubiorev.2018.09.003 |
| [40] | Bravo JA, Dinan TG, Cryan JF (2014) Early-life stress induces persistent alterations in 5-HT1A receptor and serotonin transporter mRNA expression in the adult rat brain. Front Mol Neurosci 7: 1-9. https://doi.org/10.3389/fnmol.2014.00024 |
| [41] |
González-Pardo H, Arias JL, Gómez-Lázaro E, et al. (2020) Sex-specific effects of early life stress on brain mitochondrial function, monoamine levels and neuroinflammation. Brain Sci 10: 447. https://doi.org/10.3390/brainsci10070447
|
| [42] |
Nishi M (2020) Effects of early-life stress on the brain and behaviors: implications of early maternal separation in rodents. Int J Mol Sci 21: 7212. https://doi.org/10.3390/ijms21197212
|
| [43] |
Miragaia AS, de Oliveira Wertheimer GS, Consoli AC, et al. (2018) Maternal deprivation increases anxiety-and depressive-like behaviors in an age-dependent fashion and reduces neuropeptide Y expression in the amygdala and hippocampus of male and female young adult rats. Front BehavNeurosci 12: 159. https://doi.org/10.3389/fnbeh.2018.00159
|
| [44] |
Feng P, Vurbic D, Wu Z, et al. (2007) Brain orexins and wake regulation in rats exposed to maternal deprivation. Brain Res 1154: 163-172. https://doi.org/10.1016/j.brainres.2007.03.077
|
| [45] | Marco EM, Llorente R, López-Gallardo M, et al. (2015) The maternal deprivation animal model revisited. NeurosciBiobehav Rev 51: 151-163. https://doi.org/10.1016/j.neubiorev.2015.01.015 |
| [46] |
Bülbül M, Sinen O (2022) The influence of early-life and adulthood stressors on brain neuropeptide-S system. Neuropeptides 92: 102223. https://doi.org/10.1016/j.npep.2021.102223
|
| [47] |
Harrison EL, Baune BT (2014) Modulation of early stress-induced neurobiological changes: a review of behavioural and pharmacological interventions in animal models. Transl Psychiatry 4: 390. https://doi.org/10.1038/tp.2014.31
|
| [48] |
Botticelli L, Micioni Di Bonaventura E, Ubaldi M, et al. (2021) The Neural Network of Neuropeptide S (NPS): Implications in food intake and gastrointestinal functions. Pharmaceuticals 14: 293. https://doi.org/10.3390/ph14040293
|
| [49] |
Gur E, Dremencov E, Lerer B, et al. (1999) Venlafaxine: acute and chronic effects on 5-hydroxytryptamine levels in rat brain in vivo. Eur J Pharmacol 372: 17-24. https://doi.org/10.1016/S0014-2999(99)00164-8
|
| [50] |
Dazzi L, Vignone V, Seu E, et al. (2002) Inhibition by venlafaxine of the increase in norepinephrine output in rat prefrontal cortex elicited by acute stress or by the anxiogenic drug FG 7142. J Psychopharmacol 16: 125-131. https://doi.org/10.1177/026988110201600202
|
| [51] |
Flandreau EI, Bourke CH, Ressler KJ, et al. (2013) Escitalopram alters gene expression and HPA axis reactivity in rats following chronic overexpression of corticotropin-releasing factor from the central amygdala. Psychoneuroendocrinology 38: 1349-1361. https://doi.org/10.1016/j.psyneuen.2012.11.020
|
| [52] |
Mesquita AR, Pêgo JM, Summavielle T, et al. (2007) Neurodevelopment milestone abnormalities in rats exposed to stress in early life. Neuroscience 147: 1022-1033. https://doi.org/10.1016/j.neuroscience.2007.04.007
|
| [53] |
Levine S, Huchton DM, Wiener SG, et al. (1991) Time course of the effect of maternal deprivation on the hypothalamic-pituitary-adrenal axis in the infant rat. Dev Psychobiol 24: 547-558. https://doi.org/10.1002/dev.420240803
|
| [54] |
Suchecki D, Mozaffarian D, Gross G, et al. (1993) Effects of maternal deprivation on the ACTH stress response in the infant rat. Neuroendocrinology 57: 204-212. https://doi.org/10.1159/000126361
|
| [55] | Lehmann J, Pryce CR, Bettschen D, et al. (1999) The maternal separation paradigm and adult emotionality and cognition in male and female Wistar rats. PharmacolBiochemBehav 64: 705-715. https://doi.org/10.1016/S0091-3057(99)00150-1 |
| [56] | Marcondes FK, Miguel KJ, Melo LL, et al. (2001) Estrous cycle influences the response of female rats in the elevated plus-maze test. PhysiolBehav 74: 435-440. https://doi.org/10.1016/S0031-9384(01)00593-5 |
| [57] |
Dimatelis JJ, Vermeulen IM, Bugarith K, et al. (2016) Female rats are resistant to developing the depressive phenotype induced by maternal separation stress. Metab Brain Dis 31: 109-119. https://doi.org/10.1007/s11011-015-9723-8
|
| [58] | Paxinos G, Watson C (2006) The rat brain in stereotaxic coordinates: hard cover edition. Elsevier. https://books.google.pl/books?hl=pl&lr=&id=0prYfdDbh58C&oi=fnd&pg=PP1&dq=Paxinos+G,+Watson+C.+The+rat+brain+in+stereotaxic+coordinates:+hard+cover+edition.+Elsevier.+2006.&ots=-9hrEYXGxj&sig=NOP1J-I81ZXmlNrTabF7eFjhEVM&redir_esc=y#v=onepage&q=Paxinos%20G%2C%20Watson%20C.%20The%20rat%20brain%20in%20stereotaxic%20coordinates%3A%20hard%20cover%20edition.%20Elsevier.%202006.&f=false |
| [59] |
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162: 156-159. https://doi.org/10.1016/0003-2697(87)90021-2
|
| [60] |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 25: 402-408. https://doi.org/10.1006/meth.2001.1262
|
| [61] |
Bondar NP, Merkulova TI (2016) Brain-derived neurotrophic factor and early-life stress: multifaceted interplay. J Biosci 41: 751-758. https://doi.org/10.1007/s12038-016-9648-3
|
| [62] |
Alviña K, JodeiriFarshbaf M, Mondal AK (2021) Long term effects of stress on hippocampal function: emphasis on early life stress paradigms and potential involvement of neuropeptide Y. J Neurosci Res 99: 57-66. https://doi.org/10.1002/jnr.24614
|
| [63] |
Yam KY, Ruigrok SR, Ziko I, et al. (2017) Ghrelin and hypothalamic NPY/AgRP expression in mice are affected by chronic early-life stress exposure in a sex-specific manner. Psychoneuroendocrinology 86: 73-77. https://doi.org/10.1016/j.psyneuen.2017.09.006
|
| [64] |
Guerrini R, Salvadori S, Rizzi A, et al. (2010) Neurobiology, pharmacology, and medicinal chemistry of neuropeptide S and its receptor. Med Res Rev 30: 751-777. https://doi.org/10.1002/med.20180
|
| [65] |
Okamura N, Reinscheid RK (2007) Neuropeptide S: a novel modulator of stress and arousal. Stress 10: 221-226. https://doi.org/10.1080/10253890701248673
|
| [66] |
Chauveau F, Claverie D, Lardant E, et al. (2020) Neuropeptide S promotes wakefulness through the inhibition of sleep-promoting ventrolateral preoptic nucleus neurons. Sleep 43: zsz189. https://doi.org/10.1093/sleep/zsz189
|
| [67] |
Ionescu IA, Dine J, Yen YC, et al. (2012) Intranasally administered neuropeptide S (NPS) exerts anxiolytic effects following internalization into NPS receptor-expressing neurons. Neuropsychopharmacology 37: 1323-1337. https://doi.org/10.1038/npp.2011.317
|
| [68] |
Lukas M, Bredewold R, Neumann ID, et al. (2010) Maternal separation interferes with developmental changes in brain vasopressin and oxytocin receptor binding in male rats. Neuropharmacology 58: 78-87. https://doi.org/10.1016/j.neuropharm.2009.06.020
|
| [69] |
Nauta WJ, Domesick VB (1978) Crossroads of limbic and striatal circuitry: hypothalamo-nigral connections. [In] Limbic mechanisms . Boston, MA.: Springer 75-93. https://doi.org/10.1007/978-1-4757-0716-8_6
|
| [70] |
Daniels WMU, Pietersen CY, Carstens ME, et al. (2004) Maternal separation in rats leads to anxiety-like behavior and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Metab Brain Dis 19: 3-14. https://doi.org/10.1023/B:MEBR.000027412.19664.b3
|
| [71] |
Valentino RJ, Van Bockstaele E (2001) Opposing regulation of the locus coeruleus by corticotropin-releasing factor and opioids. Psychopharmacology 158: 331-342. https://doi.org/10.1007/s002130000673
|
| [72] |
Valentino RJ, Van Bockstaele E (2008) Convergent regulation of locus coeruleus activity as an adaptive response to stress. Eur J Pharmacol 583: 194-203. https://doi.org/10.1016/j.ejphar.2007.11.062
|
| [73] |
Valentino RJ, Foote SL, Page ME (1993) The locus coeruleus as a site for integrating corticotropin-releasing factor and noradrenergic mediation of stress responses. An NY Acad Sci 697: 173-188. https://doi.org/10.1111/j.1749-6632.1993.tb49931.x
|
| [74] |
Chang S, Deussing JM (2021) Corticotropin-Releasing Hormone in the Paraventricular Nucleus of the Hypothalamus—Beyond Hypothalamic–Pituitary–Adrenal Axis Control. [In] Neuroanatomy of Neuroendocrine Systems . Cham.: Springer 231-250. https://doi.org/10.1007/978-3-030-86630-3_9
|
| [75] |
Jüngling K, Liu X, Lesting J, et al. (2012) Activation of neuropeptide S-expressing neurons in the locus coeruleus by corticotropin-releasing factor. J Physiol 590: 3701-3717. https://doi.org/10.1113/jphysiol.2011.226423
|
| [76] | Ross JA, Van Bockstaele EJ (2021) The locus coeruleus-norepinephrine system in stress and arousal: unraveling historical, current, and future perspectives. Front Psychiatry 11: 1581. https://doi.org/10.3389/fpsyt.2020.601519 |
| [77] |
Liu D, Caldji C, Sharma S, et al. (2000) Influence of neonatal rearing conditions on stress-induced adrenocorticotropin responses and norepinepherine release in the hypothalamic paraventricular nucleus. J Neuroendocrinol 12: 5-12. https://doi.org/10.1046/j.1365-2826.2000.00422.x
|
| [78] |
Matthews K, Dalley JW, Matthews C, et al. (2001) Periodic maternal separation of neonatal rats produces region-and gender-specific effects on biogenic amine content in postmortem adult brain. Synapse 40: 1-10. https://doi.org/10.1002/1098-2396(200104)40:1<1::AID-SYN1020>3.0.CO;2-E
|
| [79] |
Bhatnagar S, Vining C, Denski KAI (2004) Regulation of chronic stress-induced changes in hypothalamic-pituitary-adrenal activity by the basolateral amygdala. Ann NY Acad Sci 1032: 315-319. https://doi.org/10.1196/annals.1314.050
|
| [80] |
LeDoux J (2007) The amygdala. Curr Biol 17: 868-874. https://europepmc.org/article/med/17956742
|
| [81] |
Keshavarzi S, Sullivan RK, Ianno DJ, et al. (2014) Functional properties and projections of neurons in the medial amygdala. J Neurosci 34: 8699-8715. https://doi.org/10.1523/JNEUROSCI.1176-14.2014
|
| [82] |
Grund T, Neumann ID (2018) Neuropeptide S induces acute anxiolysis by phospholipase C-dependent signaling within the medial amygdala. Neuropsychopharmacology 43: 1156-1163. https://doi.org/10.1038/npp.2017.169
|
| [83] |
Plotsky PM, Owens MJ, Nemeroff CB (1998) Psychoneuroendocrinology of depression: hypothalamic-pituitary-adrenal axis. Psychiatr Clin N Am 21: 293-307. https://doi.org/10.1016/S0193-953X(05)70006-X
|
| [84] |
Corradi PF, Corradi RB, Greene LW (2016) Physiology of the hypothalamic pituitary gonadal axis in the male. Urol Clin 43: 151-162. https://doi.org/10.1016/j.ucl.2016.01.001
|
| [85] | Ortiga-Carvalho TM, Chiamolera MI, Pazos-Moura CC, et al. (2011) Hypothalamus-pituitary-thyroid axis. ComprPhysiol 6: 1387-1428. https://doi.org/10.1002/cphy.c150027 |
| [86] |
Veenema AH, Blume A, Niederle D, et al. (2006) Effects of early life stress on adult male aggression and hypothalamic vasopressin and serotonin. Eur J Neurosci 24: 1711-1720. https://doi.org/10.1111/j.1460-9568.2006.05045.x
|
| [87] |
Veenema AH, Neumann ID (2009) Maternal separation enhances offensive play-fighting, basal corticosterone and hypothalamic vasopressin mRNA expression in juvenile male rats. Psychoneuroendocrinology 34: 463-467. https://doi.org/10.1016/j.psyneuen.2008.10.017
|
| [88] |
Lukas M, Neumann ID (2012) Nasal application of neuropeptide S reduces anxiety and prolongs memory in rats: social versus non-social effects. Neuropharmacology 62: 398-405. https://doi.org/10.1016/j.neuropharm.2011.08.016
|
| [89] | VanBodegom M, Homberg JR, Henckens MJ (2017) Modulation of the hypothalamic-pituitary-adrenal axis by early life stress exposure. Front Cell Neurosci 11: 87. https://doi.org/10.3389/fncel.2017.00087 |
| [90] | Bombail V (2019) Perception and emotions: on the relationships between stress and olfaction. Appl BehavAnim Sci 212: 98-108. https://doi.org/10.1016/j.applanim.2018.12.013 |
| [91] |
Mikrouli E, Wörtwein G, Soylu R, et al. (2011) Increased numbers of orexin/hypocretin neurons in a genetic rat depression model. Neuropeptides 45: 401-406. https://doi.org/10.1016/j.npep.2011.07.010
|
| [92] |
Albert PR, Vahid-Ansari F (2019) The 5-HT1A receptor: signaling to behavior. Biochimie 161: 34-45. https://doi.org/10.1016/j.biochi.2018.10.015
|
| [93] |
Gilbert F, Brazell C, Tricklebank MD, et al. (1988) Activation of the 5-HT1A receptor subtype increases rat plasma ACTH concentration. Eur J Pharmacol 147: 431-439. https://doi.org/10.1016/0014-2999(88)90178-1
|
| [94] |
Fletcher A, Forster EA, Bill DJ, et al. (1995) Electrophysiological, biochemical, neurohormonal and behavioural studies with WAY-100635, a potent, selective and silent 5-HT1A receptor antagonist. Behav Brain Res 73: 337-353. https://doi.org/10.1016/0166-4328(96)00118-0
|
| [95] |
Valentino RJ, Foote SL, Aston-Jones G (1983) Corticotropin-releasing factor activates noradrenergic neurons of the locus coeruleus. Brain Res 270: 363-367. https://doi.org/10.1016/0006-8993(83)90615-7
|
| [96] |
Pickel VM, Joh TH, Reis DJ (1977) A serotonergic innervation of noradrenergic neurons in nucleus locus coeruleus: demonstration by immunocytochemical localization of the transmitter specific enzymes tyrosine and tryptophan hydroxylase. Brain Res 131: 197-214. https://doi.org/10.1016/0006-8993(77)90515-7
|
| [97] | Steinbusch HWM (1984) Serotonin-immunoreactive neurons and their projections in the CNS. Handbook of Chemical Neuroanatomy, Chemical Transmitter Receptors in the CNS, 3, Part II : 68-118. https://cir.nii.ac.jp/crid/1572824499186806784 |
| [98] | Leysen JE (2004) 5-HT2 receptors. CurrDrug Targets 3: 11-26. https://doi.org/10.2174/1568007043482598 |
| [99] |
Rossi DV, Burke TF, Hensler JG (2008) Differential regulation of serotonin-1A receptor-stimulated [35S] GTPγS binding in the dorsal raphe nucleus by citalopram and escitalopram. Eur J Pharmacol 583: 103-107. https://doi.org/10.1016/j.ejphar.2008.01.022
|
| [100] |
Segal M (1979) Serotonergic innervation of the locus coeruleus from the dorsal raphe and its action on responses to noxious stimuli. J Physiol 286: 401-415. https://doi.org/10.1113/jphysiol.1979.sp012628
|
| [101] |
Van Bockstaele EJ (2000) Multiple substrates for serotonergic modulation of rat locus coeruleus neurons and relationships with kainate receptors. Brain Res Bull 51: 433-442. https://doi.org/10.1016/S0361-9230(99)00267-1
|
| [102] |
West CH, Ritchie JC, Boss-Williams KA, et al. (2009) Antidepressant drugs with differing pharmacological actions decrease activity of locus coeruleus neurons. Int J Neuropsychopharmacol 12: 627-641. https://doi.org/10.1017/S1461145708009474
|
| [103] | Bobker DH, Williams JT (1989) Serotonin agonists inhibit synaptic potentials in the rat locus ceruleus in vitro via 5-hydroxytryptamine1A and 5-hydroxytryptamine1B receptors. J Pharmacol Exp Ther 250: 37-43. https://europepmc.org/article/med/2526217 |
| [104] |
Holanda VA, Oliveira MC, Souza LS, et al. (2019) Dopamine D1 and D2 receptors mediate neuropeptide S-induced antinociception in the mouse formalin test. Eur J Pharmacol 859: 172557. https://doi.org/10.1016/j.ejphar.2019.172557
|
| [105] |
Nitz D, Siegel J (1997) GABA release in the dorsal raphe nucleus: role in the control of REM sleep. Am J PhysiolRegulIntegr Comp Physiol 273: 451-455. https://doi.org/10.1152/ajpregu.1997.273.1.R451
|
| [106] |
Soya S, Sakurai T (2020) Orexin as a modulator of fear-related behavior: hypothalamic control of noradrenaline circuit. Brain Res 1731: 146037. https://doi.org/10.1016/j.brainres.2018.11.032
|
| [107] |
Tobinski AM, Rappeneau V (2021) Role of the Neuropeptide S System in Emotionality, Stress Responsiveness and Addiction-Like Behaviours in Rodents: Relevance to Stress-Related Disorders. Pharmaceuticals 14: 780. https://doi.org/10.3390/ph14080780
|
| [108] |
Tillage RP, Foster SL, Lustberg D, et al. (2021) Co-released norepinephrine and galanin act on different timescales to promote stress-induced anxiety-like behavior. Neuropsychopharmacology 46: 1535-1543. https://doi.org/10.1038/s41386-021-01011-8
|
| [109] | Millan MJ, Dekeyne A, Papp M, et al. (2001) S33005, a novel ligand at both serotonin and norepinephrine transporters: II. Behavioral profile in comparison with venlafaxine, reboxetine, citalopram, and clomipramine. J Pharmacol Exp Ther 298: 581-591. https://jpet.aspetjournals.org/content/298/2/565.short |
| [110] |
Rossi DV, Valdez M, Gould GG, et al. (2006) Chronic administration of venlafaxine fails to attenuate 5-HT1A receptor function at the level of receptor-G protein interaction. Int J Neuropsychopharmacol 9: 393-406. https://doi.org/10.1017/S1461145705005754
|
| [111] |
Béïque JC, Blier P, de Montigny C, et al. (2000) Potentiation by (-) pindolol of the activation of postsynaptic 5-HT1A receptors induced by venlafaxine. Neuropsychopharmacology 23: 294-306. https://doi.org/10.1016/S0893-133X(00)00112-3
|
| [112] |
Niimi M (2006) Centrally administered neuropeptide S activates orexin-containing neurons in the hypothalamus and stimulates feeding in rats. Endocrine 30: 75-79. https://doi.org/10.1385/ENDO:30:1:75
|
| [113] |
Cannella N, Kallupi M, Ruggeri B, et al. (2013) The role of the neuropeptide S system in addiction: focus on its interaction with the CRF and hypocretin/orexin neurotransmission. Prog Neurobiol 100: 48-59. https://doi.org/10.1016/j.pneurobio.2012.09.005
|
| [114] |
Berlau DJ, McGaugh JL (2006) Enhancement of extinction memory consolidation: the role of the noradrenergic and GABAergic systems within the basolateral amygdala. Neurobiol Learn Mem 86: 123-132. https://doi.org/10.1016/j.nlm.2005.12.008
|
| [115] |
Roozendaal B, Castello NA, Vedana G, et al. (2008) Noradrenergic activation of the basolateral amygdala modulates consolidation of object recognition memory. Neurobiol Learn Mem 90: 576-579. https://doi.org/10.1016/j.nlm.2008.06.010
|
| [116] |
Han RW, Xu HJ, Wang P, et al. (2014) Neuropeptide S interacts with the basolateral amygdala noradrenergic system in facilitating object recognition memory consolidation. Neurobiol Learn Mem 107: 32-36. https://doi.org/10.1016/j.nlm.2013.10.010
|
| [117] |
Guarnieri LO, Pereira-Caixeta AR, Medeiros DC, et al. (2020) Pro-neurogenic effect of fluoxetine in the olfactory bulb is concomitant to improvements in social memory and depressive-like behavior of socially isolated mice. Transl Psychiatry 10: 1-14. https://doi.org/10.1038/s41398-020-0701-5
|
| [118] | Zhang J, Dennis KA, Darling RD, et al. (2013) Neonatal citalopram exposure decreases serotonergic fiber density in the olfactory bulb of male but not female adult rats. Front Cell Neurosci 7: 67. https://doi.org/10.3389/fncel.2013.00067 |
| [119] |
Thase ME, Shelton RC, Khan A (2006) Treatment with venlafaxine extended release after SSRI nonresponse or intolerance: a randomized comparison of standard-and higher-dosing strategies. J Clin Psychopharmacol 26: 250-258. https://doi.org/10.1097/01.jcp.0000219922.19305.08
|
| [120] |
Engelmann J, Wagner S, Solheid A, et al. (2021) Tolerability of high-dose venlafaxine after switch from escitalopram in nonresponding patients with major depressive disorder. J Clin Psychopharmacol 41: 62-66. https://doi.org/10.1097/JCP.0000000000001312
|
| [121] | Bonapersona V, Kentrop J, Van Lissa CJ, et al. (2019) The behavioral phenotype of early life adversity: A 3-level meta-analysis of rodent studies. NeurosciBiobehav Rev 102: 299-307. |
| [122] |
Bonapersona V, Hoijtink H, Sarabdjitsingh RA, et al. (2021) Increasing the statistical power of animal experiments with historical control data. Nat Neurosci 24: 470-477. https://doi.org/10.1038/s41593-020-00792-3
|
| [123] |
Button KS, Ioannidis J, Mokrysz C, et al. (2013) Power failure: why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci 14: 365-376. https://doi.org/10.1038/nrn3475
|
 neurosci-09-03-022-s001.pdf neurosci-09-03-022-s001.pdf |
 |







Figures(8)

Miłosz Gołyszny, Michał Zieliński, Monika Paul-Samojedny, Artur Pałasz, Ewa Obuchowicz. Chronic treatment with escitalopram and venlafaxine affects the neuropeptide S pathway differently in adult Wistar rats exposed to maternal separation[J]. AIMS Neuroscience, 2022, 9(3): 395-422. doi: 10.3934/Neuroscience.2022022







 DownLoad:
DownLoad: