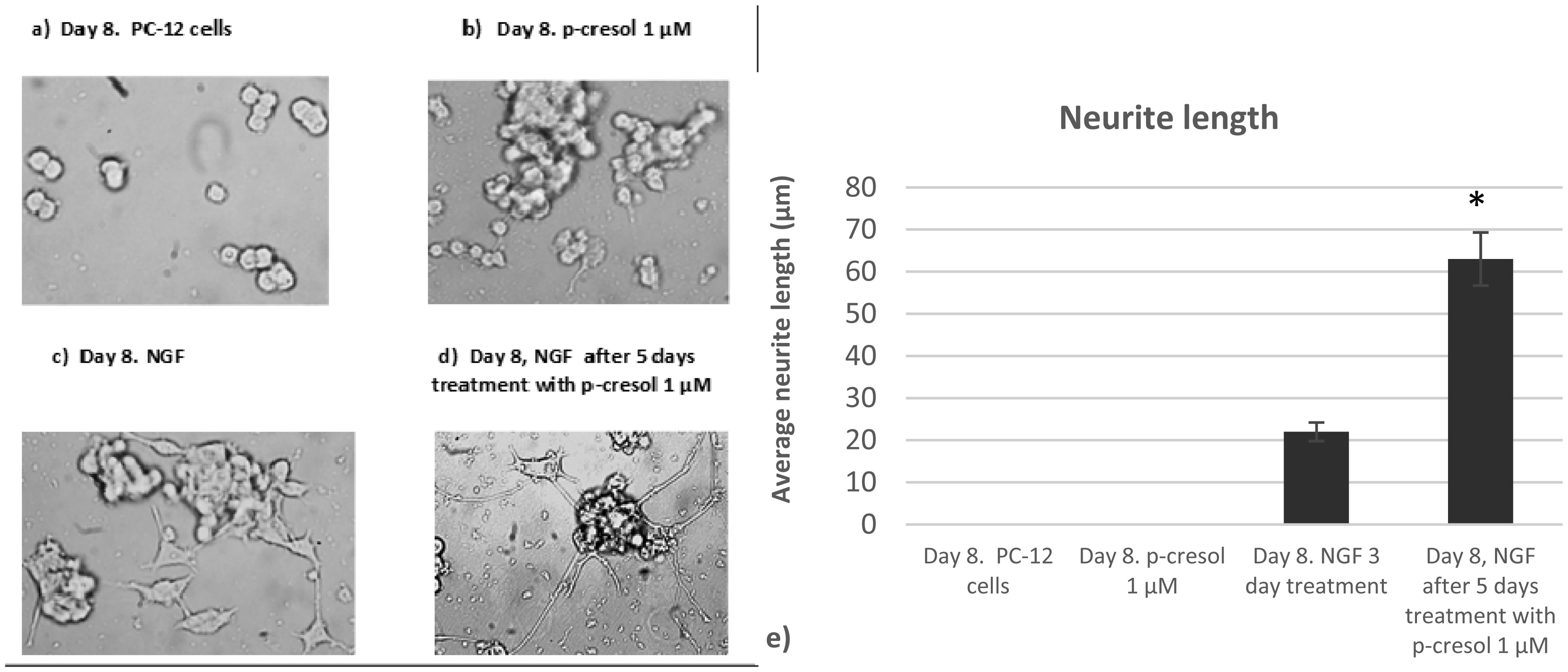
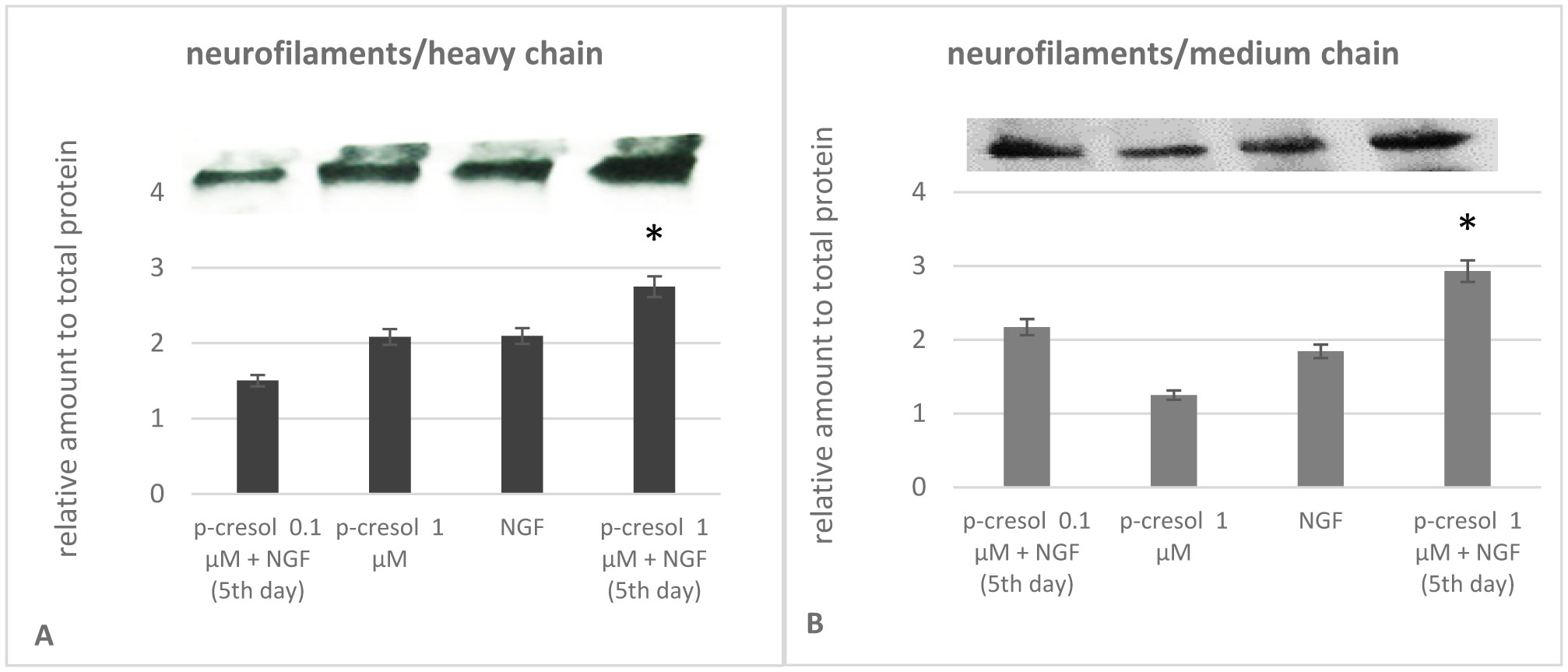
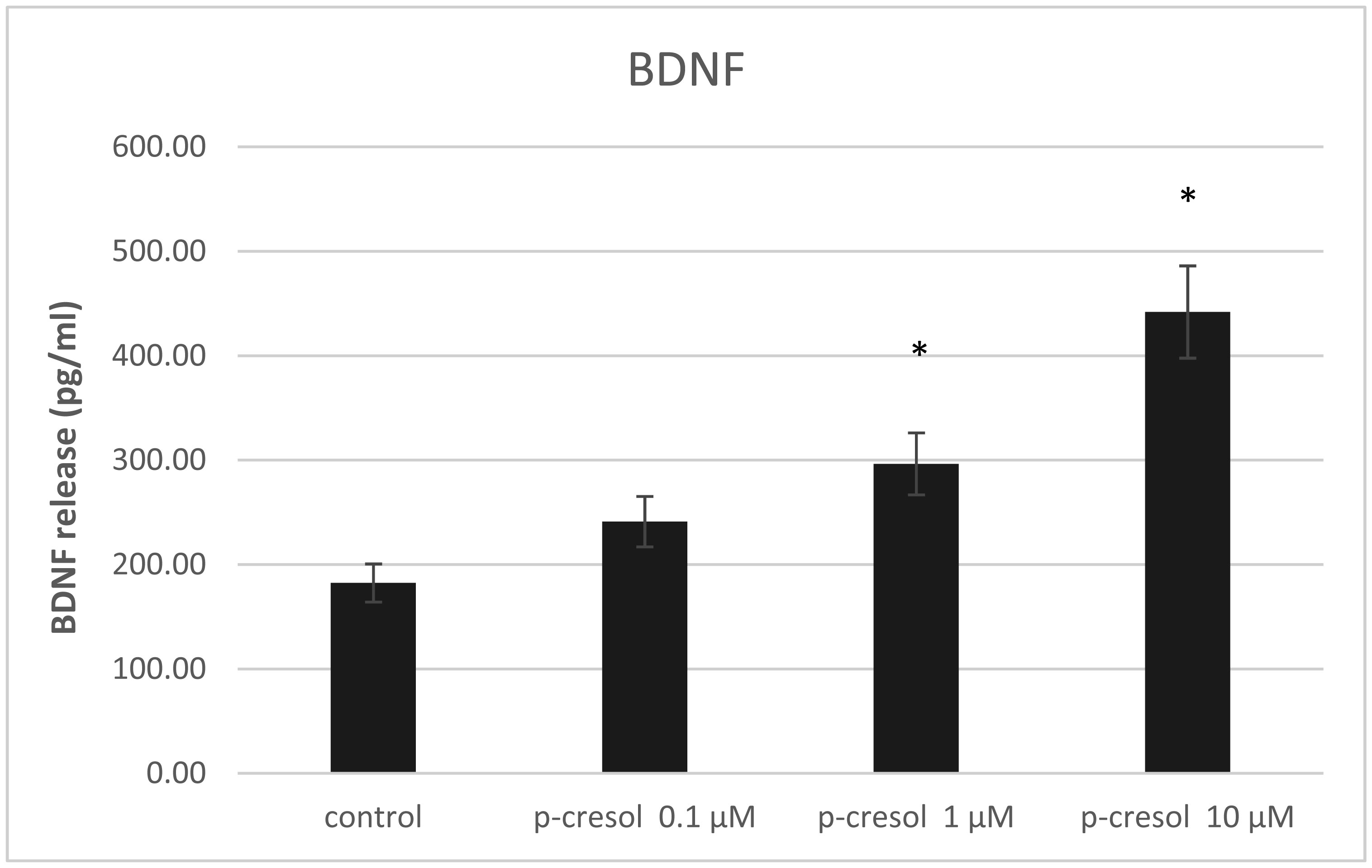
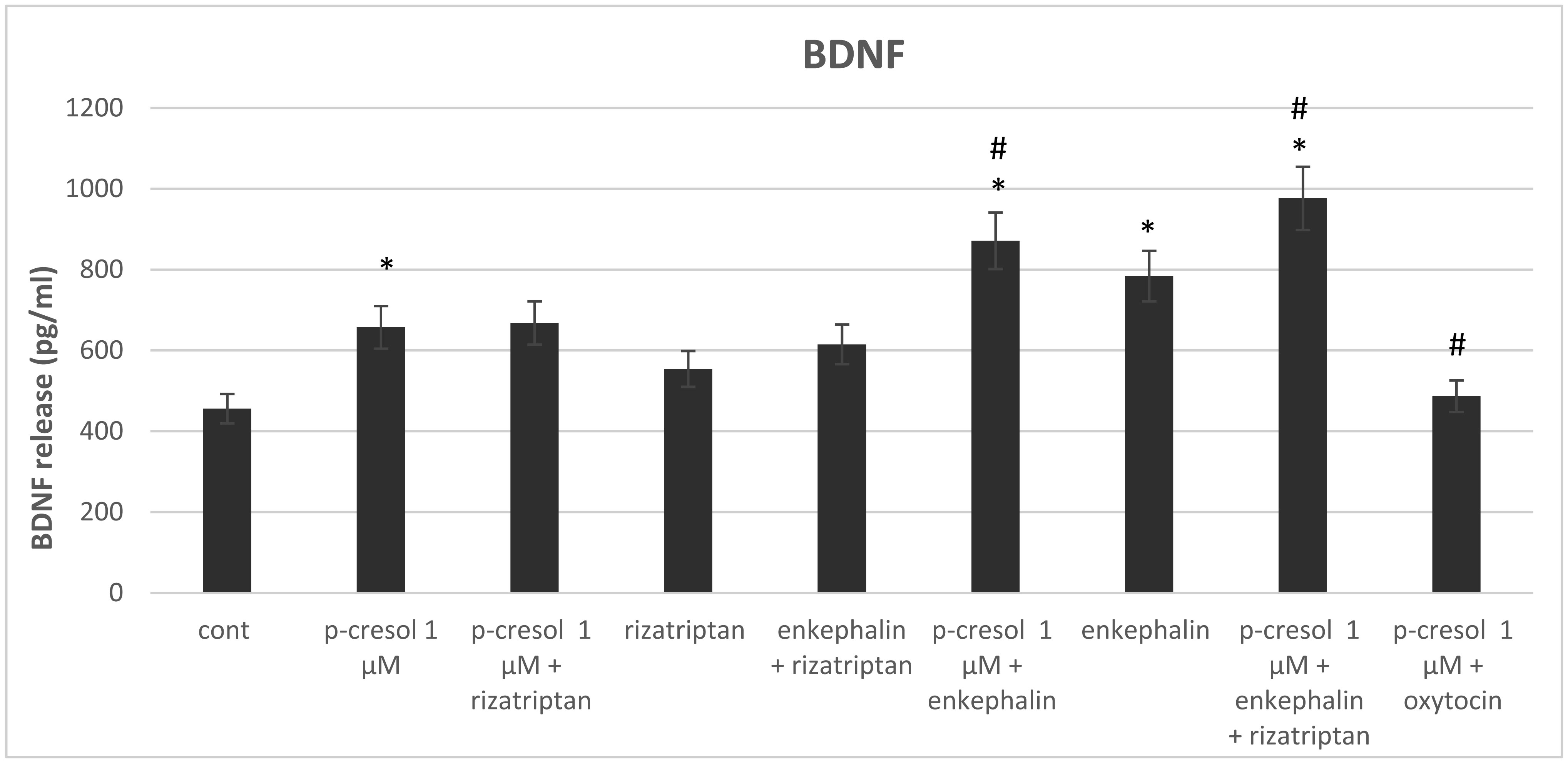
Increased p-cresol levels reportedly alter brain dopamine metabolism and exacerbate neurological disorders in experimental animals. In contrast to toxic concentrations, low doses of p-cresol may have distinct effects on neuronal metabolism. However, the role of p-cresol in synapse remodeling, neurite outgrowth, and other anabolic processes in neurons remains elusive. We propose that low doses of p-cresol affect neuronal cell structural remodeling compared with the high concentration-mediated harmful effects. Thus, the effects of p-cresol on the secretion of brain-derived neurotrophic factor (BDNF) and neurofilament subunit expression were examined using rat pheochromocytoma cells (PC-12 cells). We observed that low doses of p-cresol potentiated nerve growth factor-induced differentiation via secretion of BDNF in cultured PC-12 cells. Opioidergic compounds modulated these p-cresol effects, which were reversed by oxytocin. We propose that this effect of p-cresol has an adaptive and compensatory character and can be attributed to the induction of oxidative stress. Accordingly, we hypothesize that low doses of p-cresol induce mild oxidative stress, stimulating BDNF release by activating redox-sensitive genes. Given that the intestinal microbiome is the primary source of endogenous p-cresol, the balance between gut microbiome strains (especially Clostridium species) and opioidergic compounds may directly influence neuroplasticity.
Citation: Gigi Tevzadze, Tamar Barbakadze, Elisabed Kvergelidze, Elene Zhuravliova, Lali Shanshiashvili, David Mikeladze. Gut neurotoxin p-cresol induces brain-derived neurotrophic factor secretion and increases the expression of neurofilament subunits in PC-12 cells[J]. AIMS Neuroscience, 2022, 9(1): 12-23. doi: 10.3934/Neuroscience.2022002
Increased p-cresol levels reportedly alter brain dopamine metabolism and exacerbate neurological disorders in experimental animals. In contrast to toxic concentrations, low doses of p-cresol may have distinct effects on neuronal metabolism. However, the role of p-cresol in synapse remodeling, neurite outgrowth, and other anabolic processes in neurons remains elusive. We propose that low doses of p-cresol affect neuronal cell structural remodeling compared with the high concentration-mediated harmful effects. Thus, the effects of p-cresol on the secretion of brain-derived neurotrophic factor (BDNF) and neurofilament subunit expression were examined using rat pheochromocytoma cells (PC-12 cells). We observed that low doses of p-cresol potentiated nerve growth factor-induced differentiation via secretion of BDNF in cultured PC-12 cells. Opioidergic compounds modulated these p-cresol effects, which were reversed by oxytocin. We propose that this effect of p-cresol has an adaptive and compensatory character and can be attributed to the induction of oxidative stress. Accordingly, we hypothesize that low doses of p-cresol induce mild oxidative stress, stimulating BDNF release by activating redox-sensitive genes. Given that the intestinal microbiome is the primary source of endogenous p-cresol, the balance between gut microbiome strains (especially Clostridium species) and opioidergic compounds may directly influence neuroplasticity.

| [1] |
Hsiao EY, McBride SW, Hsien S, et al. (2013) Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 155: 1446-1448. https://doi.org/10.1016/j.cell.2013.11.024. doi: 10.1016/j.cell.2013.11.024

|
| [2] | Thakur PC, Davison JM, Stuckenholz C, et al. (2014) Dysregulated phosphatidylinositol signaling promotes endoplasmic reticulum stress-mediated intestinal mucosal injury and inflammation in zebrafish. Dis Model Mech 7: 93-106. https://doi.org/10.1242/dmm.012864. |
| [3] |
Sun CY, Li JR, Wang YY, et al. (2020) p-Cresol Sulfate Caused Behavior Disorders and Neurodegeneration in Mice with Unilateral Nephrectomy Involving Oxidative Stress and Neuroinflammation. Int J Mol Sci 21: 6687https://doi.org/10.3390/ijms21186687. doi: 10.3390/ijms21186687
|
| [4] |
Tevzadze G, Oniani N, Zhuravliova E, et al. (2019) Effects of a Gut Microbiome Toxin, p-Cresol, on the Indices of Social Behavior in Rats. Neurophysiology 50: 372-377. https://doi.org/10.1007/s11062-019-09764-1. doi: 10.1007/s11062-019-09764-1
|
| [5] |
Dawson LF, Donahue EH, Cartman ST, et al. (2011) The analysis of para-cresol production and tolerance in Clostridium difficile 027 and 012 strains. BMC Microbiol 11: 86https://doi.org/10.1186/1471-2180-11-86. doi: 10.1186/1471-2180-11-86
|
| [6] |
Bermudez-Martin P, Becker JAJ, Caramello N, et al. (2021) The microbial metabolite p-Cresol induces autistic-like behaviors in mice by remodeling the gut microbiota. Microbiome 9: 157https://doi.org/10.1186/s40168-021-01103-z. doi: 10.1186/s40168-021-01103-z
|
| [7] |
Pascucci T, Colamartino M, Fiori E, et al. (2020) P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse. Brain Sci 10: 233https://doi.org/10.3390/brainsci10040233. doi: 10.3390/brainsci10040233
|
| [8] |
Southan C, DeWolf WE, Kruse LI (1990) Inactivation of dopamine β-hydroxylase by p-cresol: Evidence for a second, minor site of covalent modification at tyrosine 357. Biochim Biophys Acta 1037: 256-258. https://doi.org/10.1016/0167-4838(90)90176-G. doi: 10.1016/0167-4838(90)90176-G
|
| [9] |
Pavǎl D (2017) A dopamine hypothesis of autism spectrum disorder. Dev Neurosci 39: 355-360. https://doi.org/10.1016/10.1159/000478725. doi: 10.1159/000478725
|
| [10] |
Tevzadze G, Zhuravliova E, Barbakadze T, et al. (2020) Gut neurotoxin p-cresol induces differential expression of GLUN2B and GLUN2A subunits of the NMDA receptor in the hippocampus and nucleus accumbens in healthy and audiogenic seizure-prone rats. AIMS Neurosci 7: 30-42. https://doi.org/10.1016/10.3934/Neuroscience.2020003. doi: 10.3934/Neuroscience.2020003
|
| [11] |
Tevzadze G, Nanobashvili Z, Zhuravliova E, et al. (2019) Effects of a Gut Microbiome Toxin, p-Cresol, on the Susceptibility to Seizures in Rats. Neurophysiology 50: 424-427. https://doi.org/10.1007/s11062-019-09774-z. doi: 10.1007/s11062-019-09774-z
|
| [12] |
Michael H, Lindenbaum S, Salvatore Carbonettot, et al. (1988) Transcriptional and Post-transcriptional Effects of Nerve Growth Factor on Expression of the Three Neurofilament Subunits in PC-12 Cells. J Biol Chem 263: 5662-5667. https://doi.org/10.1016/S0021-9258(18)60616-7. doi: 10.1016/S0021-9258(18)60616-7
|
| [13] |
Greene LA, Tischler AS (1976) Establishment of a noradrenergic clonal line of rat adrenal pheochromocytomacells which respond to nerve growth factor. Proc Natl Acad Sci USA 73: 2424-2428. https://doi.org/10.1073/pnas.73.7.2424. doi: 10.1073/pnas.73.7.2424
|
| [14] |
Wang H, Yuan G, Prabhakar NR, et al. (2006) Secretion of brain-derived neurotrophic factor from PC12 cells in response to oxidative stress requires autocrine dopamine signalling. J Neurochem 96: 694-705. https://doi.org/10.1111/j.1471-4159.2005.03572.x. doi: 10.1111/j.1471-4159.2005.03572.x
|
| [15] |
Ogura Y, Kazunori Sato, Ken-Ichi Kawashima, et al. (2014) Subtoxic levels of hydrogen peroxide induce brain-derived neurotrophic factor expression to protect PC12 cells. BMC Research Notes 7: 840https://doi.org/10.1186/1756-0500-7-840. doi: 10.1186/1756-0500-7-840
|
| [16] |
Chang MC, Chang HH, Chan CP, et al. (2014) p-Cresol affects reactive oxygen species generation, cell cycle arrest, cytotoxicity and inflammation/atherosclerosis-related modulators production in endothelial cells and mononuclear cells. PLoS ONE 9: e114446https://doi.org/10.1371/journal.pone.0114446. doi: 10.1371/journal.pone.0114446
|
| [17] |
Sheng S, Huang J, Ren Y, et al. (2018) Neuroprotection against hypoxic/ischemic injury: δ-opioid receptors and BDNF-TrkB pathway. Cell Physiol Biochem 47: 302-315. https://doi.org/10.1159/000489808. doi: 10.1159/000489808
|
| [18] |
Vanholder R, De Smet R, Lesaffer G (1999) p-Cresol: a toxin revealing many neglected but relevant aspects of uraemic toxicity. Nephrol Dial Transplant 14: 2813-2815. doi: 10.1093/ndt/14.12.2813
|
| [19] |
Hung LW, Neuner S, Polepalli JS, et al. (2017) Gating of social reward by oxytocin in the ventral tegmental area. Science 357: 1406-11. https://doi.org/10.1126/science.aan4994. doi: 10.1126/science.aan4994
|
| [20] |
Kumar H, Lim HW, More SV, et al. (2012) The role of free radicals in the aging brain and Parkinson's disease: convergence and parallelism. Int J Mol Sci 13: 10478-10504. https://doi.org/10.3390/ijms130810478. doi: 10.3390/ijms130810478
|
| [21] |
Doorn J, Florang V, Schamp J, et al. (2014) Aldehyde dehydrogenase inhibition generates a reactive dopamine metabolite autotoxic to dopamine neurons. Parkinsonism Relat Ds 20: S73-S75. https://doi.org/10.1016/S1353-8020(13)70019-1. doi: 10.1016/S1353-8020(13)70019-1
|
| [22] |
DeWolf WE, Carr SA, Varrichio A, et al. (1988) Inactivation of dopamine beta-hydroxylase by p-cresol: isolation and characterization of covalently modified active site peptides. Biochemistry 27: 9093-101. https://doi.org/10.1021/bi00426a005. doi: 10.1021/bi00426a005
|
| [23] |
Liem RK, Yen SH, Salomon GD, et al. (1978) Intermediate filaments in nervous tissues. J Cell Biol 79: 637-645. https://doi.org/10.1083/jcb.79.3.637. doi: 10.1083/jcb.79.3.637
|
| [24] |
Yuan A, Sershen H, Veeranna V, et al. (2015) Neurofilament subunits are integral components of synapses and modulate neurotransmission and behavior in vivo. Mol Psychiatry 20: 986-994. https://doi.org/10.1038/mp.2015.45. doi: 10.1038/mp.2015.45
|
| [25] |
Kim OJ, Ariano MA, Lazzarini RA, et al. (2002) Neurofilament-M interacts with the D1 dopamine receptor to regulate cell surface expression and desensitization. J Neurosci 22: 5920-5930. https://doi.org/10.1523/JNEUROSCI.22-14-05920.2002. doi: 10.1523/JNEUROSCI.22-14-05920.2002
|
| [26] | Li C, Wang H, Wang M, et al. (2021) Oxytocin attenuates methamphetamine-induced apoptosis via oxytocin receptor in rat hippocampal neurons. Front Pharmacol 12: 2063https://doi.org/10.3389/fphar.2021.639571. |
| [27] |
Martinez JL, Janak PH, Weinberger SB, et al. (1990) Enkephalin influences on behavioral and neural plasticity: mechanisms of action. NIDA Res Monogr 97: 48-78. In: |
| [28] |
Bruel-Jungerman E, Davis S, Laroche S (2007) Brain Plasticity Mechanisms and Memory: A Party of Four. Neuroscientist 13: 492-505. https://doi.org/10.1177/1073858407302725. doi: 10.1177/1073858407302725
|
| [29] |
Mahncke HW, Bronstone A, Merzenich MM (2006) Brain plasticity and functional losses in the aged: scientific bases for a novel intervention. Prog Brain Res 157: 81-109. https://doi.org/10.1016/S0079-6123(06)57006-2. doi: 10.1016/S0079-6123(06)57006-2
|
| [30] |
Moeller AH, Li Y, Mpoudi Ngole E, et al. (2014) Rapid changes in the gut microbiome during human evolution. P Natl Acad Sci 111: 16431-16435. https://doi.org/10.1073/pnas.1419136111. doi: 10.1073/pnas.1419136111
|
Figures(4)

Gigi Tevzadze, Tamar Barbakadze, Elisabed Kvergelidze, Elene Zhuravliova, Lali Shanshiashvili, David Mikeladze. Gut neurotoxin p-cresol induces brain-derived neurotrophic factor secretion and increases the expression of neurofilament subunits in PC-12 cells[J]. AIMS Neuroscience, 2022, 9(1): 12-23. doi: 10.3934/Neuroscience.2022002







 DownLoad:
DownLoad: