Citation: Wei Huo, Xiao-Min Zhu, Xin-Yan Pan, Min Du, Zhuo Sun, Zhi-Min Li. MicroRNA-527 inhibits TGF-β/SMAD induced epithelial-mesenchymal transition via downregulating SULF2 expression in non-small-cell lung cancer[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4607-4621. doi: 10.3934/mbe.2019231

| [1] | J. Su, J. Liao, L. Gao, et al., Analysis of small nucleolar RNAs in sputum for lung cancer diagnosis, Oncotarget, 7 (2016), 5131–5142. |
| [2] | L. A. Torre, R. L. Siegel and A. Jemal, Lung cancer statistics, Adv. Exp. Med. Biol., 893 (2016), 1–19. |
| [3] | L. K. Hou, Y. S. Ma, Y. Han, et al., Association of microRNA-33a molecular signature with non-small cell lung cancer diagnosis and prognosis after chemotherapy, PLoS One, 12 (2017), e0170431. |
| [4] | B. C. Roy, T. Kohno, R. Iwakawa, et al., Involvement of LKB1 in epithelial-mesenchymal transition (EMT) of human lung cancer cells, Lung Cancer, 70 (2010), 136–145. |
| [5] | B. Salehi, E. M. Varoni, M. Sharifi-Rad, et al., Epithelial-mesenchymal transition as a target for botanicals in cancer metastasis, Phytomedicine, 55 (2019), 125–136. |
| [6] | Y. Ke, W. Zhao, J. Xiong, et al., miR-149 inhibits non-small-cell lung cancer cells emt by targeting FOXM1, Biochem. Res. Int., 2013 (2013), 506731. |
| [7] | P. Nasarre, R. M. Gemmill, V. A. Potiron, et al., Neuropilin-2 Is upregulated in lung cancer cells during TGF-β1-induced epithelial-mesenchymal transition, Cancer Res., 73 (2013), 7111–7121. |
| [8] | L. Wang, X. Tong, Z. Zhou, et al., Circular RNA hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced epithelial-mesenchymal transition and metastasis by controlling TIF1gamma in non-small cell lung cancer, Mol. Cancer, 17 (2018), 140. |
| [9] | Y. Xiang, Y. Zhang, Y. Tang, et al., MALAT1 modulates TGF-β1-induced endothelial-to-mesenchymal transition through downregulation of miR-145, Cell Physiol. Biochem., 42 (2017), 357–372. |
| [10] | L. Fang, S. Wu, X. Zhu, et al., MYEOV functions as an amplified competing endogenous RNA in promoting metastasis by activating TGF-β pathway in NSCLC, Oncogene, 38 (2019), 896–912. |
| [11] | M. Tessema, C. M. Yingling, C. L. Thomas, et al., SULF2 methylation is prognostic for lung cancer survival and increases sensitivity to topoisomerase-I inhibitors via induction of ISG15, Oncogene, 31 (2012), 4107–4116. |
| [12] | G. Chen, I. Nakamura, R. Dhanasekaran, et al., Transcriptional induction of periostin by a sulfatase 2-tgf-β1-smad signaling axis mediates tumor angiogenesis in hepatocellular carcinoma, Cancer Res., 77 (2017), 632–645. |
| [13] | M. Tantawy, M. G. Elzayat, D. Yehia, et al., Identification of microRNA signature in different pediatric brain tumors, Genet. Mol. Biol., 41 (2018), 27–34. |
| [14] | 14. L. Rodriguez Calleja, C. Jacques, F. Lamoureux, et al., ΔNp63α silences a miRNA program to aberrantly initiate a wound-healing program that promotes TGF-β-induced metastasis, Cancer Res., 76 (2016), 3236–3251. |
| [15] | A. P. Mishra, B. Salehi, M. Sharifi-Rad, et al., Programmed cell death, from a cancer perspective: An overview, Mol. Diagn. Ther., 22 (2018), 281–295. |
| [16] | B. Salehi, P. Zucca, M. Sharifi-Rad, et al., Phytotherapeutics in cancer invasion and metastasis, Phytother Res., 32 (2018), 1425–1449. |
| [17] | C. Lu, H. Wang, S. Chen, et al., Baicalein inhibits cell growth and increases cisplatin sensitivity of A549 and H460 cells via miR-424-3p and targeting PTEN/PI3K/Akt pathway, J. Cell Mol. Med., 22 (2018), 2478–2487. |
| [18] | M. Zhang, C. Gao, Y. Yang, et al., MiR-424 Promotes non-small cell lung cancer progression and metastasis through regulating the tumor suppressor gene TNFAIP1, Cell Physiol. Biochem., 42 (2017), 211–221. |
| [19] | A. Izzotti, G. A. Calin, P. Arrigo, et al., Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke, Faseb J., 23 (2009), 806–812. |
| [20] | Z. Zhou, X. Niu, C. Li, et al., Inhibition of the growth of non-small cell lung cancer by miRNA-1271, Am. J. Transl. Res., 7 (2015), 1917–1924. |
| [21] | X. C. Wang, L. Q. Du, L. L. Tian, et al., Expression and function of miRNA in postoperative radiotherapy sensitive and resistant patients of non-small cell lung cancer, Lung Cancer, 72 (2011), 92–99. |
| [22] | F. K. Johansson, H. Goransson and B. Westermark, Expression analysis of genes involved in brain tumor progression driven by retroviral insertional mutagenesis in mice, Oncogene, 24 (2005), 3896–3905. |
| [23] | M. Morimoto-Tomita, K. Uchimura, A. Bistrup, et al., Sulf-2, a proangiogenic heparan sulfate endosulfatase, is upregulated in breast cancer, Neoplasia, 7 (2005), 1001–1010. |
| [24] | K. Uchimura, M. Morimoto-Tomita, A. Bistrup, et al., HSulf-2, an extracellular endoglucosamine-6-sulfatase, selectively mobilizes heparin-bound growth factors and chemokines: effects on VEGF, FGF-1, and SDF-1, BMC Biochem., 7 (2006), 2. |
| [25] | J. P. Lai, D. S. Sandhu, C. Yu, et al., Sulfatase 2 up-regulates glypican 3, promotes fibroblast growth factor signaling, and decreases survival in hepatocellular carcinoma, Hepatology, 47 (2008), 1211–1222. |
| [26] | H. Lemjabbar-Alaoui, A. van Zante, M. S. Singer, et al., Sulf-2, a heparan sulfate endosulfatase, promotes human lung carcinogenesis, Oncogene, 29 (2010), 635–646. |
| [27] | J. P. Thiery, H. Acloque, R. Y. Huang, et al., Epithelial-mesenchymal transitions in development and disease, Cell, 139 (2009), 871–890. |
| [28] | J. Yang and R. A. Weinberg, Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis, Dev. Cell, 14 (2008), 818–829. |
| [29] | J. P. Thiery, Epithelial-mesenchymal transitions in tumour progression, Nat. Rev. Cancer., 2 (2002), 442–454. |
| [30] | J. Lim and J. P. Thiery, Epithelial-mesenchymal transitions: Insights from development, Development, 139 (2012), 3471–3486. |
| [31] | D. Hanahan and R. A. Weinberg, Hallmarks of cancer: The next generation, Cell, 144 (2011), 646–674. |
| [32] | S. Lamouille, J. Xu and R. Derynck, Molecular mechanisms of epithelial-mesenchymal transition, Nat. Rev. Mol. Cell Biol., 15 (2014), 178–196. |
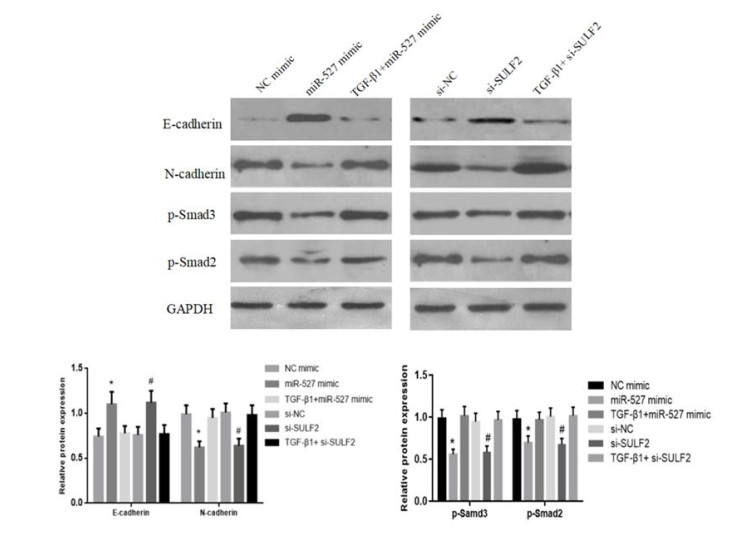
Figures(5)

Wei Huo, Xiao-Min Zhu, Xin-Yan Pan, Min Du, Zhuo Sun, Zhi-Min Li. MicroRNA-527 inhibits TGF-β/SMAD induced epithelial-mesenchymal transition via downregulating SULF2 expression in non-small-cell lung cancer[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4607-4621. doi: 10.3934/mbe.2019231







 DownLoad:
DownLoad: