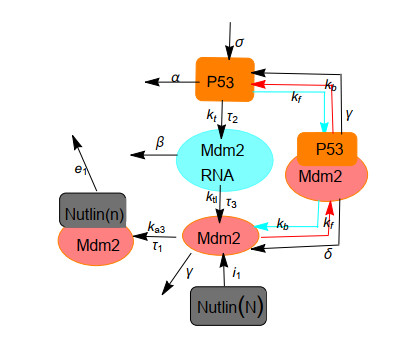
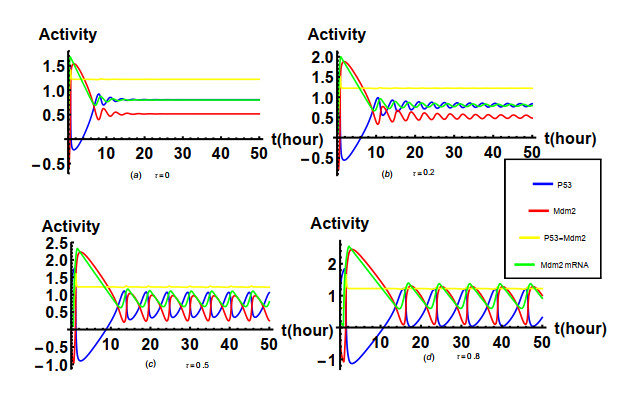
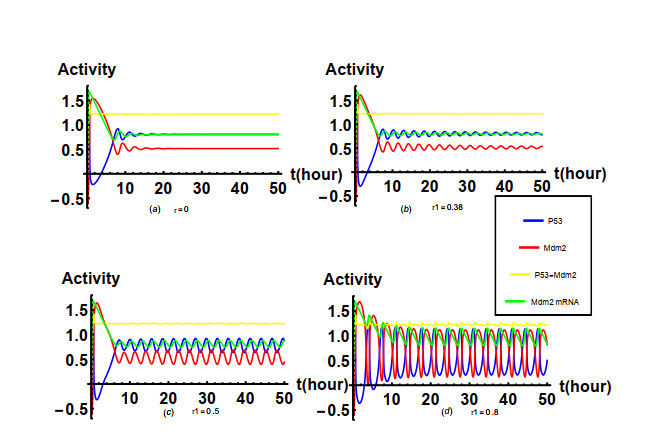
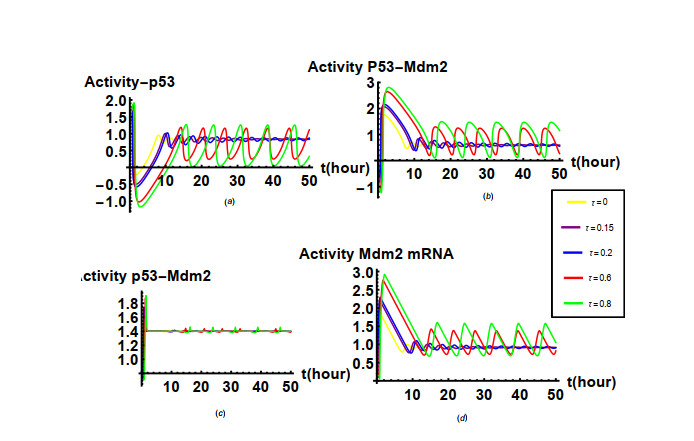
Nutlin is a family of p53-targeting drugs. It is able to bind to Mdm2, thereby accelerate the accumulation of p53 that is a prominent tumor suppressor. An integrated module of the Nutlin PBK and p53 pathway is composed of positive feedback mediated by Mdm2 mRNA as well as the drug Nutlin and negative feedback mediated by Mdm2 protein. The main research content of our paper is how the time delay of protein synthesis, response time delay of Nutlin drug, the degradation rate of Mdm2, the degradation rate of p53 depended on Mdm2 and the actual dose of Nutlin in the cell affect the oscillatory behavior caused by Hopf bifurcation in the integrated network system of Nutlin PBK and p53 pathways. The stability of the unique positive equilibrium point and the existence of Hopf bifurcation are studied by taking the time delays as the bifurcation parameters and applying bifurcation theory. Based on the normal form theory and central manifold theorem, explicit criteria to determine the Hopf bifurcation direction and stability of the bifurcated periodic solution are established. It is found that the time delays and key parameters in the integrated network system of Nutlin PBK and p53 pathways play an important role in the amplitude and period of p53 oscillation according to the results from the numerical simulation and theoretical calculation. These results may provide us with a better understanding of the biological functions of the p53 pathway and some clues for cancer treatment.
Citation: Juenu Yang, Fang Yan, Haihong Liu. Dynamic behavior of the p53-Mdm2 core module under the action of drug Nutlin and dual delays[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3448-3468. doi: 10.3934/mbe.2021173
Nutlin is a family of p53-targeting drugs. It is able to bind to Mdm2, thereby accelerate the accumulation of p53 that is a prominent tumor suppressor. An integrated module of the Nutlin PBK and p53 pathway is composed of positive feedback mediated by Mdm2 mRNA as well as the drug Nutlin and negative feedback mediated by Mdm2 protein. The main research content of our paper is how the time delay of protein synthesis, response time delay of Nutlin drug, the degradation rate of Mdm2, the degradation rate of p53 depended on Mdm2 and the actual dose of Nutlin in the cell affect the oscillatory behavior caused by Hopf bifurcation in the integrated network system of Nutlin PBK and p53 pathways. The stability of the unique positive equilibrium point and the existence of Hopf bifurcation are studied by taking the time delays as the bifurcation parameters and applying bifurcation theory. Based on the normal form theory and central manifold theorem, explicit criteria to determine the Hopf bifurcation direction and stability of the bifurcated periodic solution are established. It is found that the time delays and key parameters in the integrated network system of Nutlin PBK and p53 pathways play an important role in the amplitude and period of p53 oscillation according to the results from the numerical simulation and theoretical calculation. These results may provide us with a better understanding of the biological functions of the p53 pathway and some clues for cancer treatment.

| [1] |
J. K. Kim, T. L. Jackson, Mechanisms that enhance sustainability of p53 pulses, Plos One, 8 (2013), e65242. doi: 10.1371/journal.pone.0065242

|
| [2] |
A. Hunziker, M. H. Jensen, S. Krishna, Stress-specific response of the p53-Mdm2 feedback loop, BMC Syst. Biol., 4 (2010), 94. doi: 10.1186/1752-0509-4-94
|
| [3] | R. B. Zhao, K. Gish, M. Murphy, Y. X. Yin, A. J. Levine, Analysis of p53-regulated gene expression patterns using oligonucleotide arrays, Gene. Dev., 14 (2010), 981–993. |
| [4] |
M. Haseeb, S. Azam, A. I. Bhatti, R. Azam, M. Ullah, S. Fazal, On p53 revival using system oriented drug dosage design, J. Theor. Biol., 415 (2017), 53–57. doi: 10.1016/j.jtbi.2016.12.008
|
| [5] |
M. R. Azam, S. Fazal, M. Ullah, A.I. Bhatti, System-based strategies for p53 recovery, IET Syst. Biol., 12 (2018), 101–107. doi: 10.1049/iet-syb.2017.0025
|
| [6] |
Y. Barak, T. Juven, R. Haffner, M. Oren, Mdm2 expression is induced by wild type p53 activity, EMBO J., 12 (1993), 461–468. doi: 10.1002/j.1460-2075.1993.tb05678.x
|
| [7] |
E. Drakos, A. Thomaides, L. J. Medeiros, J. Li, G. Z. Rassidakis, Inhibition of p53-murine double minute 2 interaction by nutlin-3A stabilizes p53 and induces cell cycle arrest and apoptosis in Hodgkin lymphoma, Clin. Cancer Res., 13 (2007), 3380–3387. doi: 10.1158/1078-0432.CCR-06-2581
|
| [8] |
L. C. Mulet, D. I. Serret, A. F. Santidrian, A. M. Cosialls, M. Frias, E. Castano, et al., Mdm2 antagonists activate p53 and synergize with genotoxic drugs in B-cell chronic lymphocytic leukemia cells, Blood, 107 (2006), 4109–4114. doi: 10.1182/blood-2005-08-3273
|
| [9] |
J. G. Teodoro, S. K. Evans, M. R. Green, Inhibition of tumor angiogenesis by p53: a new role for the guardian of the genome, J. Mol. Med., 85 (2007), 1175–1186. doi: 10.1007/s00109-007-0221-2
|
| [10] |
L. Ma, J. Wagner, J. J. Rice, W. Hu, A. J. Levine, G. A. Stolovitzky, A plausible model for the digital response of p53 to DNA damage, P. Natl. A, Sci. Indin B., 102 (2005), 14266–14271. doi: 10.1073/pnas.0501352102
|
| [11] |
F. Zhang, M. Tagen, S. Throm, J. Mallari, L. Miller, R. K. Guy, et al., Whole-body physiologically based pharmacokinetic model for nutlin-3a in mice after intravenous and oral administration, Drug Metab. Dispos., 39 (2011), 15–21. doi: 10.1124/dmd.110.035915
|
| [12] |
K. Puszynski, A. Gandolfi, A. D'Onofrio, The pharmacodynamics of the p53-Mdm2 targeting drug Nutlin: the role of gene-switching noise, Plos Comput. Biol., 10 (2014), e1003991. doi: 10.1371/journal.pcbi.1003991
|
| [13] | Meek, W. David, Tumour suppression by p53: a role for the DNA damage response? Nat. Rev. Cancer, 9 (2009), 714–723. |
| [14] | K. Kojima, M. Konopleva, T. Mcqueen, S. O'Brien, W. Plunkett, Mdm2 inhibitor Nutlin-3a induces p53-mediated apoptosis by transcription-dependent and transcription-independent mechanisms and may overcome Atm-mediated resistance to fludarabine in chronic lymphocytic leukemia, Blood, 108 (2006), 993–1000. |
| [15] | S. Wang, Y. J. Zhao, A. Aguilar, D. Bernard, C. Y. Yang, Targeting the Mdm2-p53 protein-protein interaction for new cancer therapy: progress and challenges, CSH Perspect. Med., 7 (2017), a026245. |
| [16] |
J. H. Ha, E. Y. Won, J. S. Shin, M. Jang, K. S. Ryu, K. H. Bae, et al., Molecular mimicry-based repositioning of nutlin-3 to anti-apoptotic Bcl-2 family proteins, J. Am. Chem. Soc., 133 (2011), 1244–1247. doi: 10.1021/ja109521f
|
| [17] |
D. Carvajal, C. Tovar, H. Yang, B. T. Vu, D. C. Heimbrook, L. T. Vassilev, Activation of p53 by MDM2 antagonists can protect proliferating cells from mitotic inhibitors, Cancer Res., 65 (2005), 1918–1924. doi: 10.1158/0008-5472.CAN-04-3576
|
| [18] |
H. Shen, C. G. Maki1, Persistent p21 expression after Nutlin-3a removal is associated with senescence-like arrest in 4N cells, J. Biol. Chem., 285 (2010), 23105–23114. doi: 10.1074/jbc.M110.124990
|
| [19] |
B. Hu, D. M. Gilkes, B. Farooqi, S. M. Sebti, J. Chen, MDMX overexpression prevents p53 activation by the MDM2 inhibitor Nutlin, J. Biol. Chem., 281 (2006), 33030–33035. doi: 10.1074/jbc.C600147200
|
| [20] |
M. Liao, C. Xu, X. Tang, Dynamical behaviors for a competition and cooperation model of enterprises with two delays, Nonlinear Dynam., 75 (2014), 257–266. doi: 10.1007/s11071-013-1063-9
|
| [21] |
J. Wagner, L. Ma, J. Rice, H. Wenwei, A. J. Levine, G. A. Stolovitzky, p53-Mdm2 loop controlled by a balance of its feedback strength and effective dampening using ATM and delayed feedback, IEE Proc.-Syst. Biol., 152 (2005), 109–118. doi: 10.1049/ip-syb:20050025
|
| [22] | W. Zhang, J. A. Fang, T. Yang, Stochastic stability of Markovian jumping genetic regulatory networks with mixed time delays, Appl. Math. Comput., 217 (2015), 7210–7225. |
| [23] |
Z. Li, H. Gao, R. K. Agarwal, Stability analysis and controller synthesis for discrete-time delayed fuzzy systems via small gain theorem, Inf. Sci., 226 (2013), 93–104. doi: 10.1016/j.ins.2012.11.008
|
| [24] | S. Shaikh, M. Reddy, S. Dhandayuthapani, K. Alagarsamy, A. Rathinavelu, Effect of Nutlin and E3 ligase inhibitor on p53 and p21 levels in hdm2 expressing lncap cells, Cancer Res., 73 (2013), 3875. |
| [25] |
Y. Harima, Y. Takashima, Y. Ueda, T. Ohtsuka, R. Kageyama, Accelerating the tempo of the segmentation clock by reducing the number of introns in the Hes7 gene, Cell Rep., 3 (2013), 1–7. doi: 10.1016/j.celrep.2012.11.012
|
| [26] |
Y. Zhang, H. H. Liu, Y. Yan, J. Zhou, Oscillatory dynamics of p38 activity with transcriptional and translational time delays, Sci. Rep., 7 (2017), 11495. doi: 10.1038/s41598-017-11149-5
|
| [27] |
A. Zajkowicz, M. Krześniak, I. Matuszczyk, M. Głowala-Kosińska, D. Butkiewicz, M. Rusin, Nutlin-3a, an MDM2 antagonist and p53 activator, helps to preserve the replicative potential of cancer cells treated with a genotoxic dose of resveratrol, Mol. Biol. Rep., 40 (2013), 5013–5026. doi: 10.1007/s11033-013-2602-7
|
| [28] | H. Hu, Z. Wang, Dynamics of Controlled Mechanical Systems with Delayed Feedback, Springer-Verlag, 1989. |
| [29] |
Y. Takashima, T. Ohtsuka, A. Gonzalez, H. Miyachi, R. Kageyama, Intronic delay is essential for oscillatory expression in the segmentation clock, P. Natl. Acad. Sci., 108 (2011), 3300–3305. doi: 10.1073/pnas.1014418108
|
| [30] |
D. Bratsun, D. Volfson, L. S. Tsimring, J. Hasty, Delay-induced Stochastic Oscillations in Gene Regulation, P. Natl. A. Sci. India B., 102 (2005), 14593–14598. doi: 10.1073/pnas.0503858102
|
| [31] |
B. Vogelstein, D. Lane, A. Levine, Surfing the p53 network, Nature, 408 (2000), 307–310. doi: 10.1038/35042675
|
| [32] | J. K. Kim, T. L. Jackson, Mechanisms that enhance sustainability of p53 pulses, Plos One, 8 (2013), 12513. |
| [33] |
J. D. Oliner, K. W. Kinzler, P. S. Meltzer, D. L. George, B. Vogelstein, Amplification of a gene encoding a p53-associated protein in human sarcomas, Nature, 358 (1992), 80–83. doi: 10.1038/358080a0
|
| [34] |
H. G. Park, S. N. Yang, K. H. Park, Regulation of p53 stability by Mdm2, Nature, 387 (1997), 299–303. doi: 10.1038/387299a0
|
| [35] |
J. H. Park, S. W. Yang, J. M. Park, S. H. Ka, J. H. Kim, Y. Y. Kong, et al., Positive feedback regulation of p53 transactivity by DNA damage-induced ISG15 modification, Nat. Commun., 7 (2016), 12513. doi: 10.1038/ncomms12513
|
| [36] |
N. A. M. Monk, Oscillatory expression of Hes1, p53, and $NF-\kappa{B}$ driven by transcriptional time delays, Curr. Biol., 13 (2003), 1409–1413. doi: 10.1016/S0960-9822(03)00494-9
|
| [37] |
J. P. Richard, Time-delay systems: an overview of some recent advances and open problems, Automatica, 39 (2003), 1667–1694. doi: 10.1016/S0005-1098(03)00167-5
|
 mbe-18-04-173-Supplementary.pdf mbe-18-04-173-Supplementary.pdf |
 |
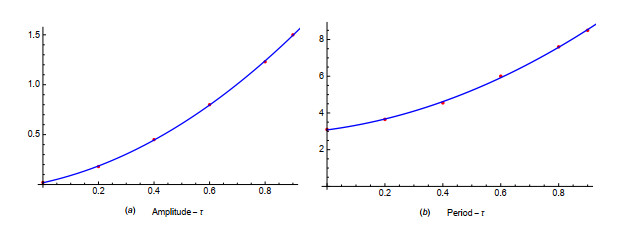
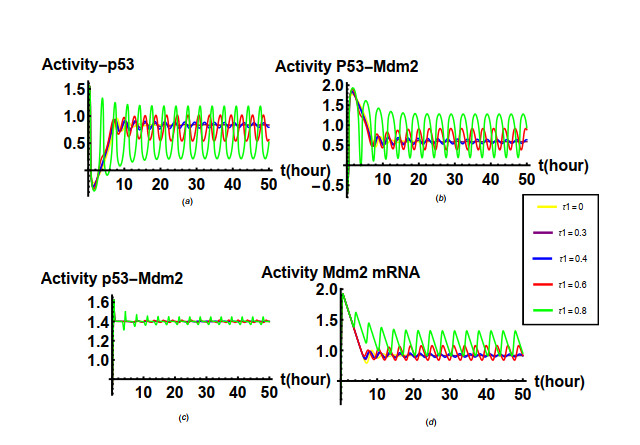
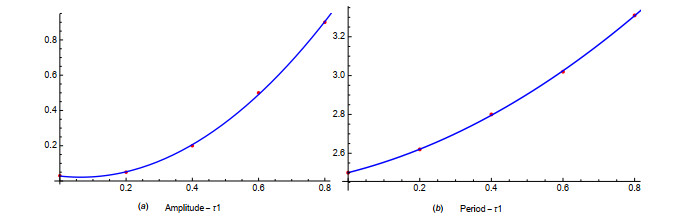
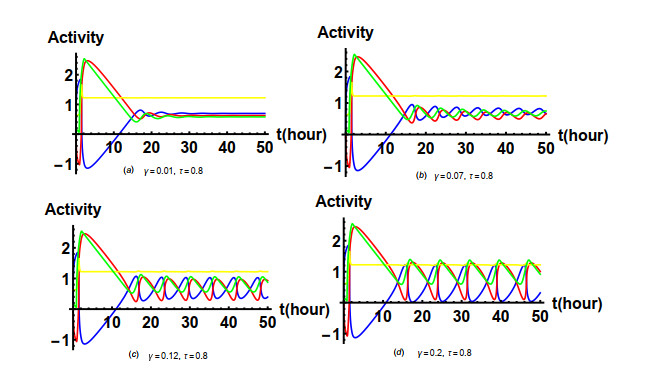
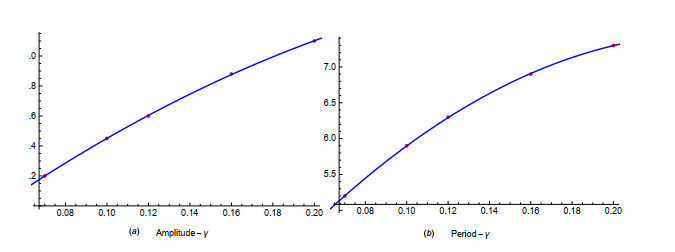
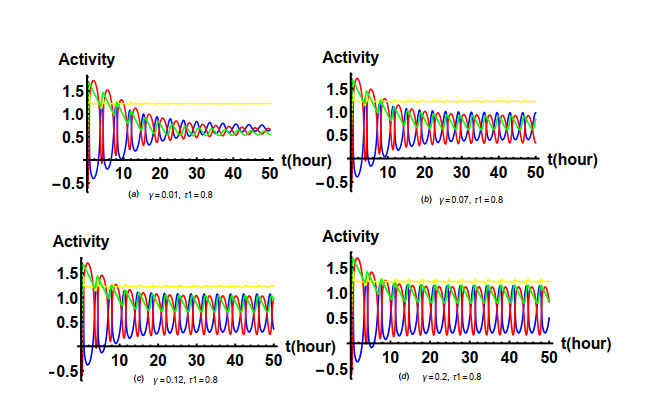
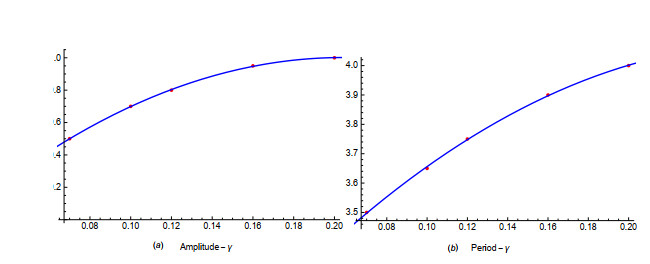
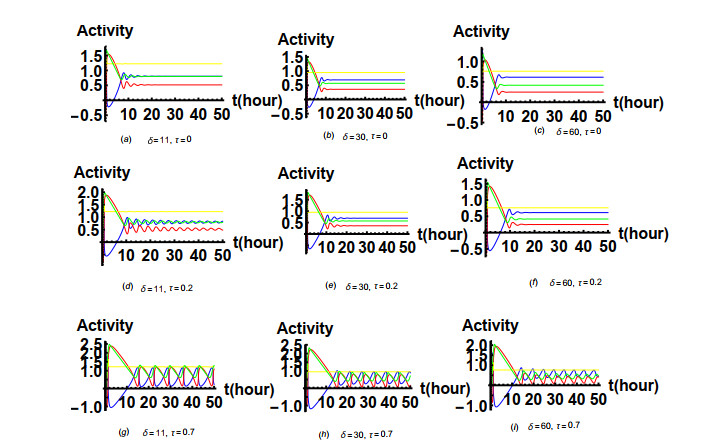
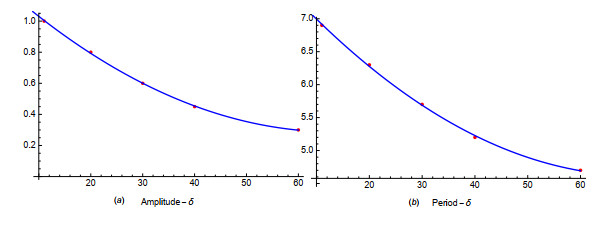
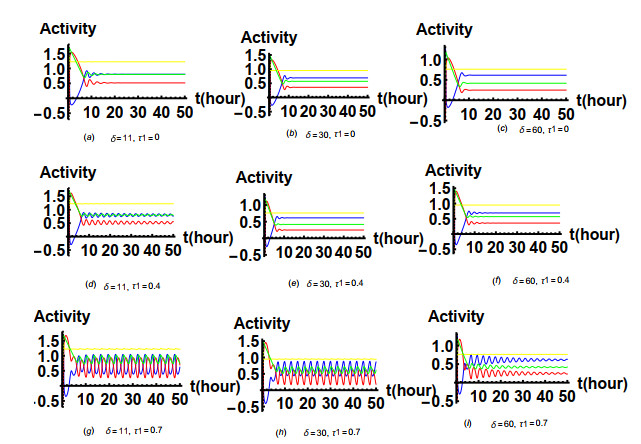
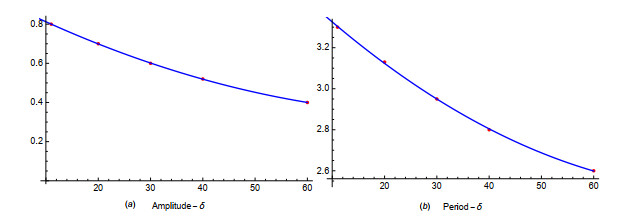
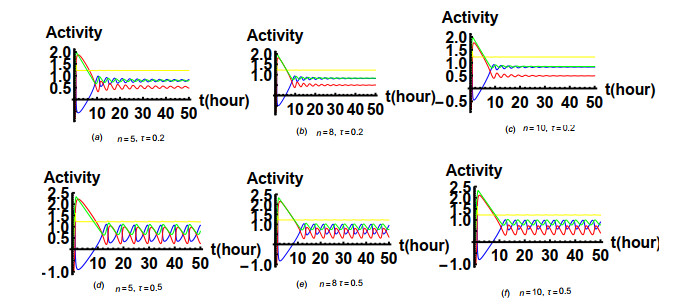
Figures(19) / Tables(1)

Juenu Yang, Fang Yan, Haihong Liu. Dynamic behavior of the p53-Mdm2 core module under the action of drug Nutlin and dual delays[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3448-3468. doi: 10.3934/mbe.2021173







 DownLoad:
DownLoad: