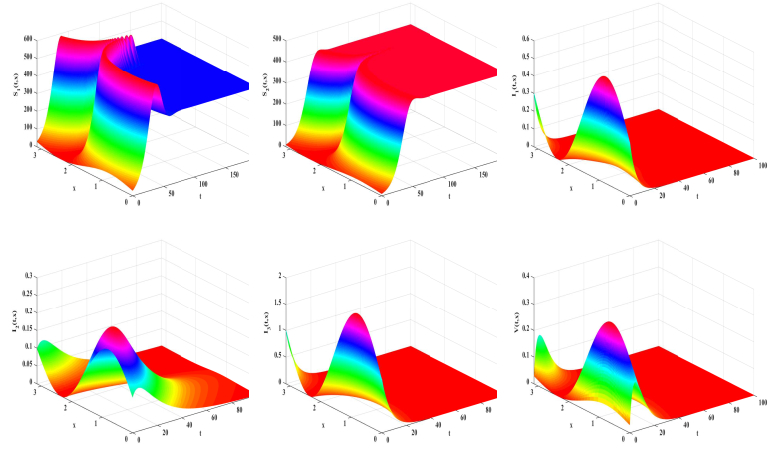
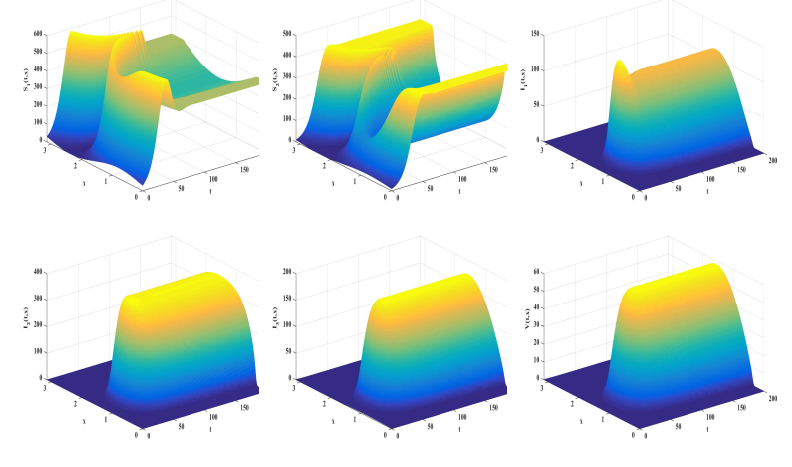
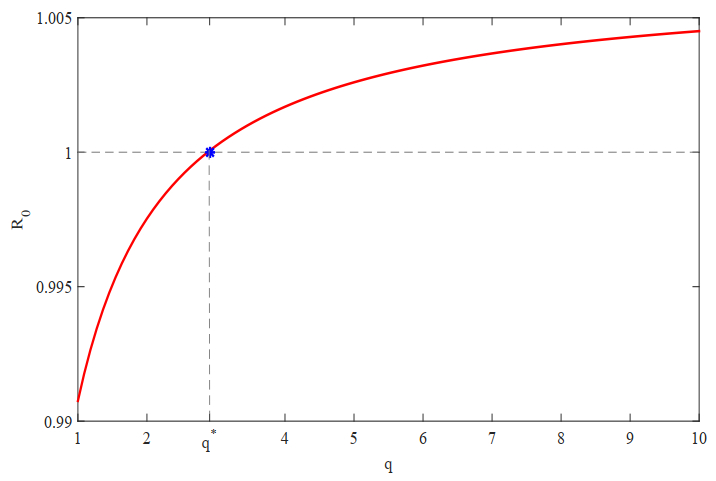
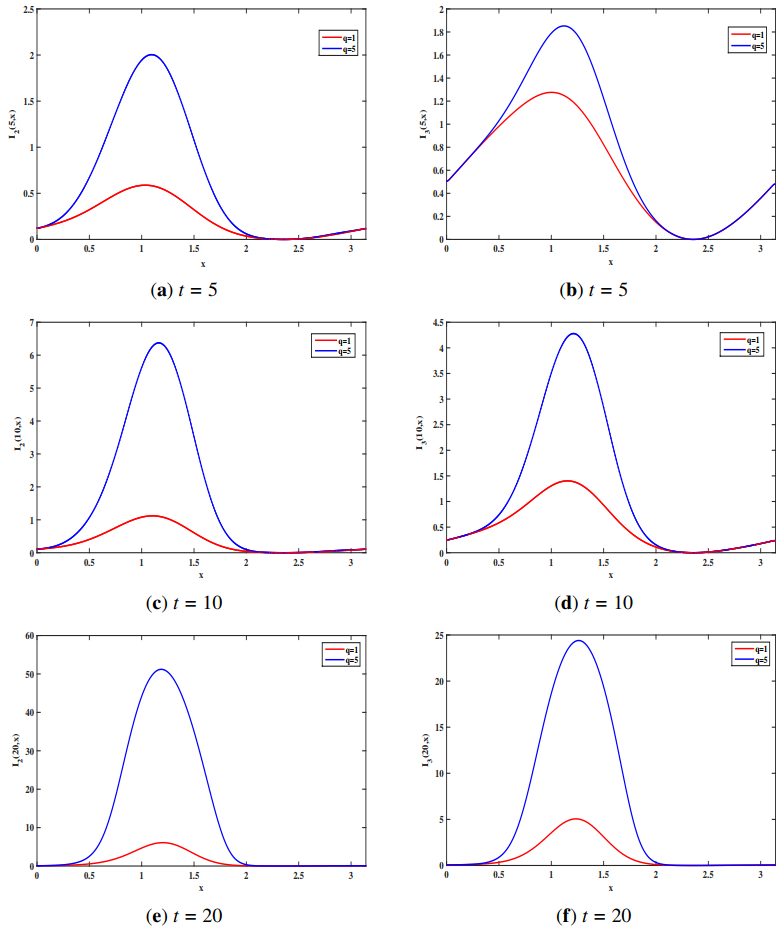
In this study, we formulate a reaction-diffusion Zika model which incorporates vector-bias, environmental transmission and spatial heterogeneity. The main question of this paper is the analysis of the threshold dynamics. For this purpose, we establish the mosquito reproduction number $ R_{1} $ and basic reproduction number $ R_{0} $. Then, we analyze the dynamical behaviors in terms of $ R_{1} $ and $ R_{0} $. Numerically, we find that the ignorance of the vector-bias effect will underestimate the infection risk of the Zika disease, ignorance of the spatial heterogeneity effect will overestimate the infection risk, and the environmental transmission is indispensable.
Citation: Liping Wang, Xinyu Wang, Dajun Liu, Xuekang Zhang, Peng Wu. Dynamical analysis of a heterogeneous spatial diffusion Zika model with vector-bias and environmental transmission[J]. Electronic Research Archive, 2024, 32(2): 1308-1332. doi: 10.3934/era.2024061
In this study, we formulate a reaction-diffusion Zika model which incorporates vector-bias, environmental transmission and spatial heterogeneity. The main question of this paper is the analysis of the threshold dynamics. For this purpose, we establish the mosquito reproduction number $ R_{1} $ and basic reproduction number $ R_{0} $. Then, we analyze the dynamical behaviors in terms of $ R_{1} $ and $ R_{0} $. Numerically, we find that the ignorance of the vector-bias effect will underestimate the infection risk of the Zika disease, ignorance of the spatial heterogeneity effect will overestimate the infection risk, and the environmental transmission is indispensable.

| [1] | World Health Organization (WHO), Emergency Committee on Zika virus and observed increase in neurological disorders and neonatal malformations, 2016. Available from: https://www.chinadaily.com.cn/world/2016-02/02/content_23348858.htm. |
| [2] |
G. Lucchese, D. Kanduc, Zika virus and autoimmunity: from microcephaly to Guillain-Barré syndrome, and beyond, Autoimmun Rev., 15 (2016), 801–808. https://doi.org/10.1016/j.autrev.2016.03.020 doi: 10.1016/j.autrev.2016.03.020

|
| [3] |
I. Bogoch, O.Brady, M. Kraemer, M. German, M. Creatore, S. Brent, et al., Potential for Zika virus introduction and transmission in resource-limited countries in Africa and the Asia-Pacific region: a modelling study, Lancet Infect Dis., 16 (2016), 1237–1245. https://doi.org/10.1016/S1473-3099(16)30270-5 doi: 10.1016/S1473-3099(16)30270-5
|
| [4] |
B. Zheng, L. Chang, J. Yu, A mosquito population replacement model consisting of two differential equations, Electron. Res. Arch., 30 (2016), 978–994. https://doi.org/10.3934/era.2022051 doi: 10.3934/era.2022051
|
| [5] |
Z. Lv, J. Zeng, Y. Ding, X. Liu, Stability analysis of time-delayed SAIR model for duration of vaccine in the context of temporary immunity for COVID-19 situation, Electron. Res. Arch., 31 (2023), 1004–1030. https://doi.org/10.3934/era.2023050 doi: 10.3934/era.2023050
|
| [6] |
H. Cao, M. Han, Y. Bai, S. Zhang, Hopf bifurcation of the age-structured SIRS model with the varying population sizes, Electron. Res. Arch., 30 (2022), 3811–3824. https://doi.org/10.3934/era.2022194 doi: 10.3934/era.2022194
|
| [7] |
X. Zhou, X. Shi, Stability analysis and backward bifurcation on an SEIQR epidemic model with nonlinear innate immunity, Electron. Res. Arch., 30 (2022), 3481–3508. https://doi.org/10.3934/era.2022178 doi: 10.3934/era.2022178
|
| [8] |
G. Fan, N. Li, Application and analysis of a model with environmental transmission in a periodic environmen, Electron. Res. Arch., 31 (2023), 5815–5844. https://doi.org/10.3934/era.2023296 doi: 10.3934/era.2023296
|
| [9] |
S. Guo, X. Yang, Z. Zheng, Global dynamics of a time-delayed malaria model with asymptomatic infections and standard incidence rate, Electron. Res. Arch., 31 (2023), 3534–3551. https://doi.org/10.3934/era.2023179 doi: 10.3934/era.2023179
|
| [10] |
K. Wang, H. Zhao, H. Wang, R. Zhang, Traveling wave of a reaction-diffusion vector-borne disease model with nonlocal effects and distributed delay, J. Dyn. Differ. Equations, 35 (2023), 3149–3185. https://doi.org/10.1007/s10884-021-10062-w doi: 10.1007/s10884-021-10062-w
|
| [11] |
K. Wang, H. Wang, H. Zhao, Aggregation and classification of spatial dynamics of vector-borne disease in advective heterogeneous environment, J. Differ. Equations, 343 (2023), 285–331. https://doi.org/10.1016/j.jde.2022.10.013 doi: 10.1016/j.jde.2022.10.013
|
| [12] |
W. E. Fitzgibbon, J. J. Morgan, G. F. Webb, An outbreak vector-host epidemic model with spatial structure: the 2015–2016 Zika outbreak in Rio De Janeiro, Theor. Biol. Med. Model., 14 (2017), 1–17. https://doi.org/10.1186/s12976-017-0051-z doi: 10.1186/s12976-017-0051-z
|
| [13] |
T. Y. Miyaoka, S. Lenhart, J. F. Meyer, Optimal control of vaccination in a vector-borne reaction-diffusion model applied to Zika virus, J. Math. Biol., 79 (2019), 1077–1104. https://doi.org/10.1007/s00285-019-01390-z doi: 10.1007/s00285-019-01390-z
|
| [14] |
K. Yamazaki, Zika virus dynamics partial differential equations model with sexual transmission route, Nonlinear Anal.-Real, 50 (2019), 290–315. https://doi.org/10.1016/j.nonrwa.2019.05.003 doi: 10.1016/j.nonrwa.2019.05.003
|
| [15] |
Y. Cai, K. Wang, W. Wang, Global transmission dynamics of a zika virus model, Appl. Math. Lett., 92 (2019), 190–195. https://doi.org/10.1016/j.aml.2019.01.015 doi: 10.1016/j.aml.2019.01.015
|
| [16] |
L. Duan, L. Huang, Threshold dynamics of a vector-host epidemic model with spatial structure and nonlinear incidence rate, Proc. Amer. Math. Soc., 149 (2021), 4789–4797. https://doi.org/10.1090/proc/15561 doi: 10.1090/proc/15561
|
| [17] |
P. Magal, G. Webb, Y. Wu, On a vector-host epidemic model with spatial structure, Nonlinearity, 31 (2018), 5589–5614. https://doi.org/10.1088/1361-6544/aae1e0 doi: 10.1088/1361-6544/aae1e0
|
| [18] |
P. Magal, G. Webb, Y. Wu, On the basic reproduction number of reaction-diffusion epidemic models, SIAM J. Appl. Math., 79 (2019), 284–304. https://doi.org/10.1137/18M1182243 doi: 10.1137/18M1182243
|
| [19] |
F. Li, X. Q. Zhao, Global dynamics of a reaction–diffusion model of Zika virus transmission with seasonality, B. Math. Biol., 83 (2021), 43. https://doi.org/10.1007/s11538-021-00879-3 doi: 10.1007/s11538-021-00879-3
|
| [20] |
S. Du, Y. Liu, J. Liu, J. Zhao, C. Champagne, L. Tong, et al., Aedes mosquitoes acquire and transmit Zika virus by breeding in contaminated aquatic environments, Nat. Commun., 10 (2019), 1324. https://doi.org/10.1038/s41467-019-09256-0 doi: 10.1038/s41467-019-09256-0
|
| [21] |
L. Wang, H. Zhao, Modeling and dynamics analysis of Zika transmission with contaminated aquatic environments, Nonlinear Dyn., 104 (2021), 845–862. https://doi.org/10.1007/s11071-021-06289-3 doi: 10.1007/s11071-021-06289-3
|
| [22] |
L. Wang, P. Wu, M. Li, L. Shi, Global dynamics analysis of a Zika transmission model with environment transmission route and spatial heterogeneity, AIMS Math., 7 (2022), 4803–4832. https://doi.org/10.3934/math.2022268 doi: 10.3934/math.2022268
|
| [23] |
L. Wang, P. Wu, Threshold dynamics of a Zika model with environmental and sexual transmissions and spatial heterogeneity, Z. Angew. Math. Phys., 73 (2022), 1–22. https://doi.org/10.1007/s00033-022-01812-x doi: 10.1007/s00033-022-01812-x
|
| [24] |
J. Wang, Y. Chen, Threshold dynamics of a vector-borne disease model with spatial structure and vector-bias, Appl. Math. Lett., 100 (2020), 106052. https://doi.org/10.1016/j.aml.2019.106052 doi: 10.1016/j.aml.2019.106052
|
| [25] |
Y. Pan, S. Zhu, J. Wang, A note on a ZIKV epidemic model with spatial structure and vector-bias, AIMS Math., 7 (2022), 2255–2265. https://doi.org/10.3934/math.2022128 doi: 10.3934/math.2022128
|
| [26] | P. Hess, Periodic-parabolic boundary value problems and positivity, Longman Scientific and Technical, Harlow, 1991. https://doi.org/10.1112/blms/24.6.619 |
| [27] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Society, Rhode Island, 1995. https://doi.org/10.1090/surv/041 |
| [28] |
X. Ren, Y. Tian, L. Liu, X. Liu, A reaction–diffusion within-host HIV model with cell-to-cell transmission, J. Math. Biol., 76 (2018), 1831–1872. https://doi.org/10.1007/s00285-017-1202-x doi: 10.1007/s00285-017-1202-x
|
| [29] | M. Wang, Nonlinear Elliptic Equations, Science Publication, Beijing, 2010. |
| [30] |
R. H. Martin, H. L. Smith, Abstract functional-differential equations and reaction-diffusion systems, Trans. Amer. Math. Soc., 321 (1990), 1–44. https://doi.org/10.1090/S0002-9947-1990-0967316-X doi: 10.1090/S0002-9947-1990-0967316-X
|
| [31] |
W. Wang, X. Q. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Syst., 11 (2012), 1652–1673. https://doi.org/10.1137/120872942 doi: 10.1137/120872942
|
| [32] |
Y. C. Shyu, R. N. Chien, F. B. Wang, Global dynamics of a West Nile virus model in a spatially variable habitat, Nonlinear Anal.-Real, 41 (2018), 313–333. https://doi.org/10.1016/j.nonrwa.2017.10.017 doi: 10.1016/j.nonrwa.2017.10.017
|
| [33] |
S. B. Hsu, F. B. Wang, X. Q. Zhao, Global dynamics of zooplankton and harmful algae in flowing habitats, J. Differ. Equations, 255 (2013), 265–297. https://doi.org/10.1016/j.jde.2013.04.006 doi: 10.1016/j.jde.2013.04.006
|
| [34] |
S. B. Hsu, F. B. Wang, X. Q. Zhao, Dynamics of a periodically pulsed bio-reactor model with a hydraulic storage zone, J. Dyn. Differ. Equations, 23 (2011), 817–842. https://doi.org/10.1007/s10884-011-9224-3 doi: 10.1007/s10884-011-9224-3
|
| [35] |
P. Magal, X. Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM J. Math. Anal., 37 (2005), 251–275. https://doi.org/10.1137/S0036141003439173 doi: 10.1137/S0036141003439173
|
| [36] | X. Q. Zhao, Dynamical Systems in Population Biology, 2nd edition, Springer, New York, 2017. https://doi.org/10.1007/978-3-319-56433-3 |
| [37] |
R. Wu, X. Q. Zhao, A reaction-diffusion model of vector-borne disease with periodic delays, J Nonlinear Sci., 29 (2019), 29–64. https://doi.org/10.1007/s00332-018-9475-9 doi: 10.1007/s00332-018-9475-9
|
| [38] |
F. Li, X. Q. Zhao, Global dynamics of a nonlocal periodic reaction-diffusion model of bluetongue disease, J. Differ. Equations, 272 (2021), 127–163. https://doi.org/10.1016/j.jde.2020.09.019 doi: 10.1016/j.jde.2020.09.019
|
| [39] | Brazil Ministry of Health, Zika cases from the Brazil Ministry of Health, 2018. Available from: http://portalms.saude.gov.br/boletins-epidemiologicos. |
| [40] | City Populations Worldwide, Brazil population, 2018. Available from: http://population.city/brazil/. |
Figures(8)
Liping Wang, Xinyu Wang, Dajun Liu, Xuekang Zhang, Peng Wu. Dynamical analysis of a heterogeneous spatial diffusion Zika model with vector-bias and environmental transmission[J]. Electronic Research Archive, 2024, 32(2): 1308-1332. doi: 10.3934/era.2024061







 DownLoad:
DownLoad: