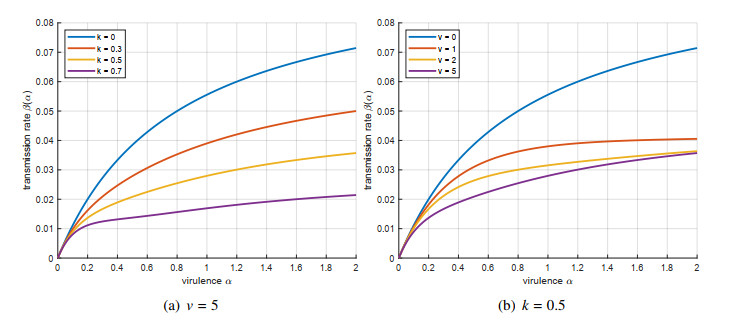
The spread of viruses can be effectively reduced by the publicity of epidemic prevention. Additionally, the interaction between heterogeneous strains has a significant effect on virus evolution. Thus, we first establish an evolutionary dynamics Susceptible-Infected- Recovered (SIR) model which considers the interaction between heterogeneous strains. We utilize adaptive dynamics to investigate the evolutionary outcomes of the trade-off between transmission and virulence. Second, we perform a critical function analysis to generalize the results independent of specific trade-off assumptions and to determine the conditions for evolutionary stability and convergence stability. Last, we investigate the effects of different publicity measures on virulence evolution under two types of interactions, including the case of excess mortality alone and the coexistence of excess mortality and superinfection. Based on the general hypothesis of transmission virulence trade-off, we introduce the cost of host mobility caused by the scope and intensity of publicity. Numerical simulations present a set of evolutionary results, including continuously stable strategies, evolutionary branching points, repellers, and the Garden of Eden. Our results indicate that an excessive publicity scope and intensity can drive the epidemic evolution towards higher virulence. Both types of interactions suggest that continuously increasing the publicity scope under a low publicity intensity can effectively reduce virulence. Furthermore, the concurrent presence of excess mortality and superinfection induces the emergence of a higher virulence.
Citation: Yike Lv, Xinzhu Meng. Evolution of infectious diseases induced by epidemic prevention publicity and interaction between heterogeneous strains[J]. Electronic Research Archive, 2024, 32(8): 4858-4886. doi: 10.3934/era.2024223
The spread of viruses can be effectively reduced by the publicity of epidemic prevention. Additionally, the interaction between heterogeneous strains has a significant effect on virus evolution. Thus, we first establish an evolutionary dynamics Susceptible-Infected- Recovered (SIR) model which considers the interaction between heterogeneous strains. We utilize adaptive dynamics to investigate the evolutionary outcomes of the trade-off between transmission and virulence. Second, we perform a critical function analysis to generalize the results independent of specific trade-off assumptions and to determine the conditions for evolutionary stability and convergence stability. Last, we investigate the effects of different publicity measures on virulence evolution under two types of interactions, including the case of excess mortality alone and the coexistence of excess mortality and superinfection. Based on the general hypothesis of transmission virulence trade-off, we introduce the cost of host mobility caused by the scope and intensity of publicity. Numerical simulations present a set of evolutionary results, including continuously stable strategies, evolutionary branching points, repellers, and the Garden of Eden. Our results indicate that an excessive publicity scope and intensity can drive the epidemic evolution towards higher virulence. Both types of interactions suggest that continuously increasing the publicity scope under a low publicity intensity can effectively reduce virulence. Furthermore, the concurrent presence of excess mortality and superinfection induces the emergence of a higher virulence.

| [1] | R. M. Anderson, R. M. May, Infectious Diseases of Humans: Dynamics and Control, Oxford university press, 1991. |
| [2] |
X. Meng, S. Zhao, W. Zhang, Adaptive dynamics analysis of a predator–prey model with selective disturbance, Appl. Math. Comput., 266 (2015), 946–958. https://doi.org/10.1016/j.amc.2015.06.020 doi: 10.1016/j.amc.2015.06.020

|
| [3] |
H. A. Adamu, M. Muhammad, A. Jingi, M. Usman, Mathematical modelling using improved sir model with more realistic assumptions, Int. J. Eng. Sci., 6 (2019), 64–69. https://doi.org/10.31873/IJEAS.6.1.22 doi: 10.31873/IJEAS.6.1.22
|
| [4] | H. H. Weiss, The SIR model and the foundations of public health, Mater. Matematics, (2013), 0001–17. |
| [5] |
L. Stone, R. Olinky, A. Huppert, Seasonal dynamics of recurrent epidemics, Nature, 446 (2007), 533–536. https://doi.org/10.1038/nature05638 doi: 10.1038/nature05638
|
| [6] | A. Pugliese, Evolutionary dynamics of virulence, Elem. Adapt. Dyn., 2000. |
| [7] | U. Dieckmann, J. Metz, M. W. Sabelis, K. Sigmund, Adaptive dynamics of infectious diseases, Pursuit Virulence Manage., (2002), 460–463. https://doi.org/10.1017/CBO9780511525728 |
| [8] | B. Boldin, S. A. H. Geritz, É. Kisdi, Superinfections and adaptive dynamics of pathogen virulence revisited: a critical function analysis, Evol. Ecol. Res., 11 (2009), 153–175. |
| [9] |
M. Wang, J. Yi, W. Jiang, Study on the virulence evolution of sars-cov-2 and the trend of the epidemics of covid-19, Math. Methods Appl. Sci., 45 (2022), 6515–6534. https://doi.org/10.1002/mma.8184 doi: 10.1002/mma.8184
|
| [10] |
E. Kisdi, S. A. H. Geritz, B. Boldin, Evolution of pathogen virulence under selective predation: a construction method to find eco-evolutionary cycles, J. Theor. Biol., 339 (2013), 140–150. https://doi.org/10.1016/j.jtbi.2013.05.023 doi: 10.1016/j.jtbi.2013.05.023
|
| [11] | E. L. Charnov, Phenotypic evolution under fisher's fundamental theorem of natural selection, Heredity, 62 (1989),, 113–116. https://doi.org/10.1038/hdy.1989.15 |
| [12] |
Roff, Trade-offs between growth and reproduction: an analysis of the quantitative genetic evidence, J. Evol. Biol., 13 (2000), 434–445. https://doi.org/10.1046/j.1420-9101.2000.00186.x doi: 10.1046/j.1420-9101.2000.00186.x
|
| [13] |
S. C. Stearns, Trade-offs in life-history evolution, Funct. Ecol., 3 (1989), 259–268. https://doi.org/10.2307/2389364 doi: 10.2307/2389364
|
| [14] |
A. Pugliese, On the evolutionary coexistence of parasite strains, Math. Biosci., 177 (2002), 355–375. https://doi.org/10.1016/S0025-5564(02)00083-4 doi: 10.1016/S0025-5564(02)00083-4
|
| [15] |
B. Boldin, O. Diekmann, Superinfections can induce evolutionarily stable coexistence of pathogens, J. Math. Biol., 56 (2008), 635–672. https://doi.org/10.1007/s00285-007-0135-1 doi: 10.1007/s00285-007-0135-1
|
| [16] |
S. Alizon, M. Baalen, Emergence of a convex trade-off between transmission and virulence, Am. Nat., 165 (2005), E155–E167. https://doi.org/10.1086/430053 doi: 10.1086/430053
|
| [17] |
S. Alizon, M. Baalen, Multiple infections, immune dynamics, and the evolution of virulence, Am. Nat., 172 (2008), E150–E168. https://doi.org/10.1086/590958 doi: 10.1086/590958
|
| [18] | R. M. May, R. M. Anderson, Epidemiology and genetics in the coevolution of parasites and hosts, in Proceedings of the Royal society of London. Series B. Biological sciences, 219 (1983), 281–313. https://doi.org/10.1098/rspb.1983.0075 |
| [19] |
A. Best, A. White, M. Boots, The implications of coevolutionary dynamics to host-parasite interactions, Am. Nat., 173 (2009), 779–791. https://doi.org/10.1086/598494 doi: 10.1086/598494
|
| [20] |
S. K. Sheppard, Strain wars and the evolution of opportunistic pathogens, Curr. Opin. Microbiol., 67 (2022), 102138. https://doi.org/10.1016/j.mib.2022.01.009 doi: 10.1016/j.mib.2022.01.009
|
| [21] |
A. J. Kucharski, V. Andreasen, J. R. Gog, Capturing the dynamics of pathogens with many strains, J. Math. Biol., 72 (2016), 1–24. https://doi.org/10.1007/s00285-015-0873-4 doi: 10.1007/s00285-015-0873-4
|
| [22] | J. R. Gog, B. T. Grenfell, Dynamics and selection of many-strain pathogens, in Proceedings of the National Academy of Sciences, 99 (2002), 17209–17214. https://doi.org/10.1073/pnas.252512799 |
| [23] |
T. Lazebnik, S. Bunimovich-Mendrazitsky, Generic approach for mathematical model of multi-strain pandemics, PloS One, 17 (2022), e0260683. https://doi.org/10.1371/journal.pone.0260683 doi: 10.1371/journal.pone.0260683
|
| [24] |
X. Cheng, Y. Wang, G. Huang, Dynamics of a competing two-strain sis epidemic model with general infection force on complex networks, Nonlinear Anal. Real World Appl., 59 (2021), 103247. https://doi.org/10.1016/j.nonrwa.2020.103247 doi: 10.1016/j.nonrwa.2020.103247
|
| [25] |
I. Gordo, M. G. M. Gomes, D. G. Reis, P. R. A. Campos, Genetic diversity in the sir model of pathogen evolution, PloS One, 4 (2009), e4876. https://doi.org/10.1371/journal.pone.0004876 doi: 10.1371/journal.pone.0004876
|
| [26] |
T. Lazebnik, Computational applications of extended sir models: A review focused on airborne pandemics, Ecol. Modell., 483 (2023), 110422. https://doi.org/10.1016/j.ecolmodel.2023.110422 doi: 10.1016/j.ecolmodel.2023.110422
|
| [27] |
E. Numfor, N. Tuncer, M. Martcheva, Optimal control of a multi-scale hiv-opioid model, J. Biol. Dyn., 18 (2024), 2317245. https://doi.org/10.1080/17513758.2024.2317245 doi: 10.1080/17513758.2024.2317245
|
| [28] |
S. C. Roberts, The evolution of hornedness in female ruminants, Behaviour, 133 (1996), 399–442. https://doi.org/10.1163/156853996X00521 doi: 10.1163/156853996X00521
|
| [29] |
E. Kisdi, Evolutionary branching under asymmetric competition, J. Theor. Biol., 197 (1999), 149–162. https://doi.org/10.1006/jtbi.1998.0864 doi: 10.1006/jtbi.1998.0864
|
| [30] |
P. V. Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [31] |
S. A. H. Geritz, E. Kisdi, G. M. NA, J. A. J. Metz, Evolutionarily singular strategies and the adaptive growth and branching of the evolutionary tree, Evol. Ecol., 12 (1998), 35–57. https://doi.org/10.1023/A:1006554906681 doi: 10.1023/A:1006554906681
|
| [32] |
J. A. J. Metz, R. M. Nisbet, S. A. H. Geritz, How should we define???fitness??? for general ecological scenarios?, Trends Ecol. Evol., 7 (1992), 198–202. https://doi.org/10.1016/0169-5347(92)90073-K doi: 10.1016/0169-5347(92)90073-K
|
| [33] |
T. L. Vincent, Y. Cohen, J. S. Brown, Evolution via strategy dynamics, Theor. Popul Biol., 44 (1993), 149–176. https://doi.org/10.1006/tpbi.1993.1023 doi: 10.1006/tpbi.1993.1023
|
| [34] | P. Jagers, Branching Processes with Biological Applications, London: Wiley, 1975. |
| [35] |
U. Dieckmann, R. Law, The dynamical theory of coevolution: a derivation from stochastic ecological processes, J. Math. Biol., 34 (1996), 579–612. https://doi.org/10.1007/BF02409751 doi: 10.1007/BF02409751
|
| [36] |
C. de Mazancourt, U. Dieckmann, Trade-off geometries and frequency-dependent selection, Am. Nat., 164 (2004), 765–778. https://doi.org/10.1086/424762 doi: 10.1086/424762
|
| [37] |
G. Meszena, E. Kisdi, U. Dieckmann, S. A. H. Geritz, J. A. J. Metz, Evolutionary optimisation models and matrix games in the unified perspective of adaptive dynamics, Selection, 2 (2002), 193–220. https://doi.org/10.1556/Select.2.2001.1-2.14 doi: 10.1556/Select.2.2001.1-2.14
|
| [38] | J. Metz, S. D. Mylius, O. Diekmann, When does evolution optimise?, IIASA Interim Report, 2008. |
| [39] |
C. Matessi, C. D. Pasquale, Long-term evolution of multilocus traits, J. Math. Biol., 34 (1996), 613–653. https://doi.org/10.1007/BF02409752 doi: 10.1007/BF02409752
|
| [40] | É. Kisdi, Trade-off geometries and the adaptive dynamics of two co-evolving species, Evol. Ecol. Res., 8 (2006), 959–973. |
| [41] |
S. Alizon, A. Hurford, N. Mideo, M. Van Baalen, Virulence evolution and the trade-off hypothesis: history, current state of affairs and the future, J. Evol. Biol., 22 (2009), 245–259. https://doi.org/10.1111/j.1420-9101.2008.01658.x doi: 10.1111/j.1420-9101.2008.01658.x
|
| [42] |
B. Boldin, E. Kisdi, On the evolutionary dynamics of pathogens with direct and environmental transmission, Evolution, 66 (2012), 2514–2527. https://doi.org/10.1111/j.1558-5646.2012.01613.x doi: 10.1111/j.1558-5646.2012.01613.x
|











Figures(12) / Tables(2)
Yike Lv, Xinzhu Meng. Evolution of infectious diseases induced by epidemic prevention publicity and interaction between heterogeneous strains[J]. Electronic Research Archive, 2024, 32(8): 4858-4886. doi: 10.3934/era.2024223







 DownLoad:
DownLoad: