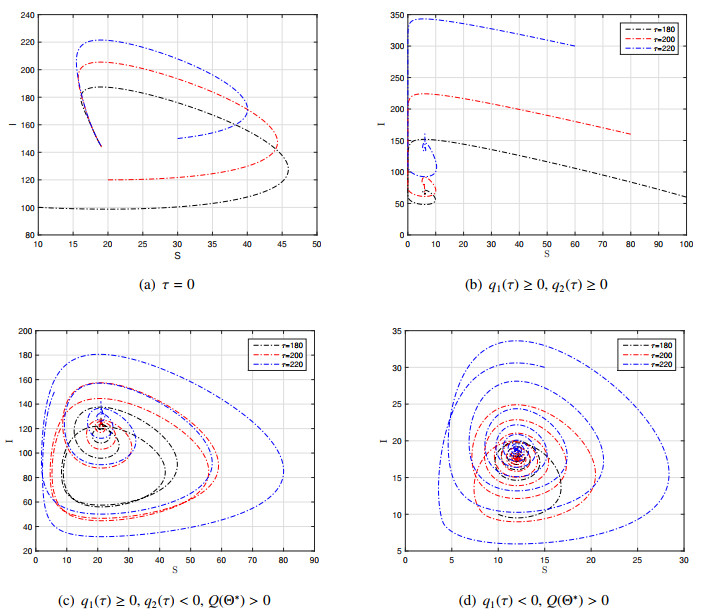
The purpose of this paper is to investigate the impact of the varying population sizes on the dynamic behavior of the age-structured epidemic model. A age-structured SIRS epidemic model with the varying population sizes is established and investigated to take into account time delay. The non-negativity of the solution, the existence and stability of the steady states, and the existence of the Hopf bifurcation are discussed. The numerical simulations show that the varying population sizes can cause the age-structured SIRS model to produce multiple stability switches.
Citation: Hui Cao, Mengmeng Han, Yunxiao Bai, Suxia Zhang. Hopf bifurcation of the age-structured SIRS model with the varying population sizes[J]. Electronic Research Archive, 2022, 30(10): 3811-3824. doi: 10.3934/era.2022194
The purpose of this paper is to investigate the impact of the varying population sizes on the dynamic behavior of the age-structured epidemic model. A age-structured SIRS epidemic model with the varying population sizes is established and investigated to take into account time delay. The non-negativity of the solution, the existence and stability of the steady states, and the existence of the Hopf bifurcation are discussed. The numerical simulations show that the varying population sizes can cause the age-structured SIRS model to produce multiple stability switches.

| [1] |
A. Johansen, A simple model of recurrent epidemics, J. Theor. Biol., 178 (1996), 45–51. https://doi.org/10.1006/jtbi.1996.0005 doi: 10.1006/jtbi.1996.0005

|
| [2] |
M. Begon, S. Telfer, M. J. Smith, S. Burthe, S. Paterson, X. Lambin, Seasonal host dynamics drive the timing of recurrent epidemics in a wildlife population, Proc. R. Soc. B, 276 (2009), 1063–1610. https://doi.org/10.1098/rspb.2008.1732 doi: 10.1098/rspb.2008.1732
|
| [3] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. A, 115 (1927), 700–721. https://doi.org/10.1098/rspa.1927.0118 doi: 10.1098/rspa.1927.0118
|
| [4] |
H. W. Hethcote, Qualitative analysis of communicable disease models, Math. Biosci., 28 (1976), 334–356. https://doi.org/10.1016/0025-5564(76)90132-2 doi: 10.1016/0025-5564(76)90132-2
|
| [5] |
J. Mena-Lorca, H. W. Hethcote, Dynamic models of infectious disease as regulators of population size, J. Math. Biol., 30 (1992), 693–716. https://doi.org/10.1007/BF00173264 doi: 10.1007/BF00173264
|
| [6] |
A. Korobeinikov, Lyapunov functions and global stability for SIR and SIRS epidemiological models with nonlinear transmission, Bull. Math. Biol., 68 (2006), 615–626. https://doi.org/10.1007/s11538-005-9037-9 doi: 10.1007/s11538-005-9037-9
|
| [7] |
B. Buonomo, S. Rionero, On the Lyapunov stability for SIRS epidemic models with general nonlinear incidence rate, Appl. Math. Comput., 217 (2010), 4010–4016. https://doi.org/10.1016/j.amc.2010.10.007 doi: 10.1016/j.amc.2010.10.007
|
| [8] |
Z. Hu, P. Bi, W. Ma, S. Ruan, Bifurcations of an SIRS epidemic model with nonlinear incidence rate, Discrete Contin. Dyn. Syst., 15 (2011), 93–112. https://doi.org/10.3934/dcdsb.2011.15.93 doi: 10.3934/dcdsb.2011.15.93
|
| [9] |
Y. Muroya, H. Li, T. Kuniya, Complete global analysis of an SIRS epidemic model with graded cure and incomplete recovery rates, J. Math. Anal. Appl., 410 (2014), 719–732. https://doi.org/10.1016/j.jmaa.2013.08.024 doi: 10.1016/j.jmaa.2013.08.024
|
| [10] |
Q. Liu, Q. Chen, Analysis of the deterministic and stochastic SIRS epidemic models with nonlinear incidence, Physica A, 428 (2015), 140–153. https://doi.org/10.1016/j.physa.2015.01.075 doi: 10.1016/j.physa.2015.01.075
|
| [11] |
T. Li, F. Zhang, H. Liu, Y. Chen, Threshold dynamics of an SIRS model with nonlinear incidence rate and transfer from infectious to susceptible, Appl. Math. Lett., 70 (2017), 52–57. https://doi.org/10.1016/j.aml.2017.03.005 doi: 10.1016/j.aml.2017.03.005
|
| [12] |
G. Lan, S. Yuan, B. Song, The impact of hospital resources and environmental perturbations to the dynamics of SIRS model, J. Franklin Inst., 358 (2021), 2405–2433. https://doi.org/10.1016/j.jfranklin.2021.01.015 doi: 10.1016/j.jfranklin.2021.01.015
|
| [13] |
M. Levine, R. Black, M. Clements, L. Cisneros, D. R. Nalin, C. R. Young, Duration of infection-derived immunity to cholera, J. Infect. Dis., 143 (1981), 818–820. https://doi.org/10.1093/infdis/143.6.818 doi: 10.1093/infdis/143.6.818
|
| [14] |
H. J. Wearing, P. Rohani, Estimating the duration of pertussis immunity using epidemiological signatures, PLoS Pathog., 5 (2009), e1000647. https://doi.org/10.1371/journal.ppat.1000647 doi: 10.1371/journal.ppat.1000647
|
| [15] |
S. Gupta, R. W. Snow, C. A. Donnelly, K. Marsh, C. Newbold, Immunity to non-cerebral severe malaria is acquired after one or two infections, Nat. Med., 5 (1999), 340–343. https://doi.org/10.1038/6560 doi: 10.1038/6560
|
| [16] |
B. F. Finkenst$\ddot{a}$dt, O. N. Bj$\phi$rnstad, B. T. Grenfell, A stochastic model for extinction and recurrence of epidemics: estimation and inference for measles outbreaks, Biostatistics, 3 (2002), 493–510. https://doi.org/10.1093/biostatistics/3.4.493 doi: 10.1093/biostatistics/3.4.493
|
| [17] |
X. C. Duan, J. F. Yin, X. Z. Li, Global Hopf bifurcation of an SIRS epidemicmodel with age-dependent recovery, Chaos, Solitons Fractals, 104 (2017), 613–624. https://doi.org/10.1016/j.chaos.2017.09.029 doi: 10.1016/j.chaos.2017.09.029
|
| [18] |
H. Cao, D. Yan, A. Li, Dynamic analysis of the recurrent epidemic model, Math. Biosci. Eng., 16 (2019), 5972–5990. https://doi.org/10.3934/mbe.2019299 doi: 10.3934/mbe.2019299
|
| [19] |
H. Cao, X. Gao, J. Li, D. Yan, Z. Yue, The bifurcation analysis of an SIRS epidemic model with immunity age and constant treatment, Appl. Anal., 100 (2021), 2844–2866. https://doi.org/10.1080/00036811.2019.1698728 doi: 10.1080/00036811.2019.1698728
|
| [20] |
T. Yu, T. Zhang, S. Yuan, Dynamic analysis of a size-structured model describing insect destruction in forests, Chaos, Solitons Fractals, 159 (2022), 112116. https://doi.org/10.1016/j.chaos.2022.112116 doi: 10.1016/j.chaos.2022.112116
|
| [21] |
E. Beretta, Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144–1165. https://doi.org/10.1137/S0036141000376086 doi: 10.1137/S0036141000376086
|
| [22] | J. K. Hale, Theory of Function Differential Equations, Springer, Heidelberg, 1977. |




Figures(5)
Hui Cao, Mengmeng Han, Yunxiao Bai, Suxia Zhang. Hopf bifurcation of the age-structured SIRS model with the varying population sizes[J]. Electronic Research Archive, 2022, 30(10): 3811-3824. doi: 10.3934/era.2022194







 DownLoad:
DownLoad: