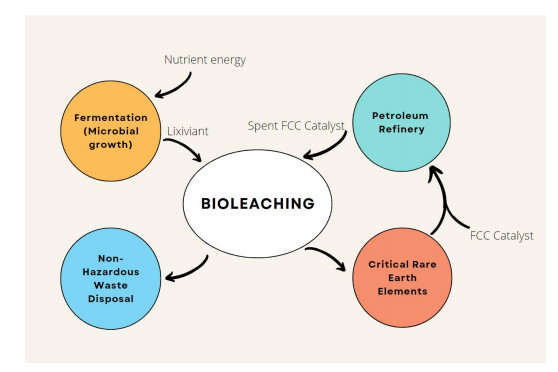
Critical metals, which mainly include the platinum group of metals, and the rare earth elements, have gained much importance because these elements are essential for economic development. A matter of concern is their availability, which is scarce, and so a constant supply is at risk. Bioleaching is one of the commonly used methods to extract these critical metals from various sources, such as industrial wastewater and mining water.
In this study, we have discussed the mechanisms of bioleaching, the factors that affect bioleaching, and a correlation between the extraction of the critical metals using microalgae which has many positive aspects. The review also suggests the future prospects for the use of microalgae in the extraction of critical metals.
Citation: Susmita Mukherjee, Sharanya Paul, Shreya Bhattacharjee, Somava Nath, Upasana Sharma, Sonali Paul. Bioleaching of critical metals using microalgae[J]. AIMS Environmental Science, 2023, 10(2): 226-244. doi: 10.3934/environsci.2023013
Critical metals, which mainly include the platinum group of metals, and the rare earth elements, have gained much importance because these elements are essential for economic development. A matter of concern is their availability, which is scarce, and so a constant supply is at risk. Bioleaching is one of the commonly used methods to extract these critical metals from various sources, such as industrial wastewater and mining water.
In this study, we have discussed the mechanisms of bioleaching, the factors that affect bioleaching, and a correlation between the extraction of the critical metals using microalgae which has many positive aspects. The review also suggests the future prospects for the use of microalgae in the extraction of critical metals.

| [1] |
Hagelüken C (2014) Recycling of (critical) metals. Critical metals handbook 2014: 41–69. https://doi.org/10.1002/9781118755341.ch3 doi: 10.1002/9781118755341.ch3

|
| [2] |
Zhuang WQ, Fitts JP, Ajo-Franklin CM, et al. (2015) Recovery of critical metals using biometallurgy. Current opinion in biotechnology 33: 327–335. https://doi.org/10.1016/j.copbio.2015.03.019 doi: 10.1016/j.copbio.2015.03.019
|
| [3] | Ayres RU, Peiro LT (2013) Material Efficiency: rare and critical metals. Philosophical transactions of the royal society A-mathematical physical and engineering sciences: 371: 20110563-569.2011 Impact factor: 2.773. Philosophical transactions of the royal society A-mathematical physical and engineering sciences 371: 563–596. https://doi.org/10.1098/rsta.2011.0563 |
| [4] |
Fu F, Wang Q (2011) Removal of heavy metal ions from wastewaters: a review. Journal of environmental management 92: 407–418. https://doi.org/10.1016/j.jenvman.2010.11.011 doi: 10.1016/j.jenvman.2010.11.011
|
| [5] |
Ongondo FO, Williams ID, Cherrett TJ (2011) How are WEEE doing? A global review of the management of electrical and electronic wastes. Waste Management 31: 714–730. https://doi.org/10.1016/j.wasman.2010.10.023 doi: 10.1016/j.wasman.2010.10.023
|
| [6] | US National Research Council, Committee on Critical Mineral Impacts of the U.S. (2008) Economy, Committee on Earth Resources. Minerals, critical minerals, and the U.S economy. Washington, DC: The National Academies Press. |
| [7] | Rath B (2012) Microalgal bioremediation: current practices and perspectives. Journal of Biochemical Technology 3: 299–304. |
| [8] |
Perales-Vela HV, Peña-Castro JM, Canizares-Villanueva RO (2006) Heavy metal detoxification in eukaryotic microalgae. Chemosphere 64: 1–10. https://doi.org/10.1016/j.chemosphere.2005.11.024 doi: 10.1016/j.chemosphere.2005.11.024
|
| [9] | Monteiro CM, Castro PM, Malcata FX (2011) Microalga-mediated bioremediation of heavy metal–contaminated surface waters. In Biomanagement of metal-contaminated soils 2011: 365–385. https://doi.org/10.1007/978-94-007-1914-9_16 |
| [10] |
Binnemans K, Jones PT, Blanpain B, et al. (2013) Recycling of rare earths: a critical review. Journal of cleaner production 51: 1–22. https://doi.org/10.1016/j.jclepro.2012.12.037. doi: 10.1016/j.jclepro.2012.12.037
|
| [11] |
Tunsu C, Petranikova M, Gergorić M, et al. (2015) Reclaiming rare earth elements from end-of-life products: A review of the perspectives for urban mining using hydrometallurgical unit operations. Hydrometallurgy 156: 239–258. https://doi.org/10.1016/j.hydromet.2015.06.007. doi: 10.1016/j.hydromet.2015.06.007
|
| [12] |
Graedel TE, Harper EM, Nassar NT, et al. (2015) On the materials basis of modern society. Proceedings of the National Academy of Sciences 112: 6295–6300. https://doi.org/10.1073/pnas.1312752110. doi: 10.1073/pnas.1312752110
|
| [13] |
Deplanche K, Macaskie LE (2008) Bio recovery of gold by Escherichia coli and Desulfovibrio desulfuricans. Biotechnology and bioengineering 99: 1055–1064. https://doi.org/10.1002/bit.21688 doi: 10.1002/bit.21688
|
| [14] |
Prabhu S, Poulose EK (2012) Silver nanoparticles: mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. International nano letters 2: 1–10. https://doi.org/10.1186/2228-5326-2-32 doi: 10.1186/2228-5326-2-32
|
| [15] |
B Bai, F Jia, Y Yang, et al. (2019) Multiple physiological response analyses aid the understanding of sensitivity variation between Microcystis aeruginosa and Chlorella sp. under paraquat exposures. Environmental Sciences Europe 31: 1–17. https://doi.org/10.1186/s12302-019-0255-4 doi: 10.1186/s12302-019-0255-4
|
| [16] |
Rempel A, Gutkoski JP, Nazari MT, et al. (2021) Current advances in microalgae-based bioremediation and other technologies for emerging contaminants treatment. The Science of the total environment 772: 144918. https://doi.org/10.1016/j.scitotenv.2020.144918 doi: 10.1016/j.scitotenv.2020.144918
|
| [17] |
Paterson-Beedle M, Readman JE, Hriljac JA, et al. (2010) Bio recovery of uranium from aqueous solutions at the expense of phytic acid. Hydrometallurgy 104: 524–528. https://doi.org/10.1016/j.hydromet.2010.01.019 doi: 10.1016/j.hydromet.2010.01.019
|
| [18] |
Kulkarni S, Ballal A, Apte SK (2013) Bioprecipitation of uranium from alkaline waste solutions using recombinant Deinococcus radiodurans. Journal of hazardous materials 262: 853–861. https://doi.org/10.1016/j.jhazmat.2013.09.057 doi: 10.1016/j.jhazmat.2013.09.057
|
| [19] |
Li Y, Wu X, Jiang X, et al. (2021) Algicidal activity of Aspergillus niger induced by calcium ion as signal molecule on Microcystis aeruginosa. Algal Research 60: 102536. https://doi.org/10.1016/j.algal.2021.102536 doi: 10.1016/j.algal.2021.102536
|
| [20] |
Marrero J, Coto O, Goldmann S, et al. (2015) Recovery of nickel and cobalt from laterite tailings by reductive dissolution under aerobic conditions using Acidithiobacillus species. Environmental science & technology 49: 6674–6682. https://doi.org/10.1021/acs.est.5b00944 doi: 10.1021/acs.est.5b00944
|
| [21] |
Capeness MJ, Edmundson MC, Horsfall LE (2015) Nickel and platinum group metal nanoparticle production by Desulfovibrio alaskensis G20. New biotechnology 32: 727–731. https://doi.org/10.1016/j.nbt.2015.02.002 doi: 10.1016/j.nbt.2015.02.002
|
| [22] |
Ashokkumar V, Chen WH, Kamyab H, et al. (2019) Cultivation of microalgae Chlorella sp. in municipal sewage for biofuel production and utilization of biochar derived from residue for the conversion of hematite iron ore (Fe2O3) to iron (Fe)–Integrated algal biorefinery. Energy 189: 116128. https://doi.org/10.1016/j.energy.2019.116128 doi: 10.1016/j.energy.2019.116128
|
| [23] |
Capeness MJ, Edmundson MC, Horsfall LE (2015) Nickel and platinum group metal nanoparticle production by Desulfovibrio alaskensis G20. New biotechnology 32: 727–731. https://doi.org/10.1016/j.nbt.2015.02.002 doi: 10.1016/j.nbt.2015.02.002
|
| [24] |
Yong P, Paterson-Beedle M, Mikheenko IP, et al. (2007) From bio-mineralisation to fuel cells: biomanufacture of Pt and Pd nanocrystals for fuel cell electrode catalyst. Biotechnology letters 29: 539–544. https://doi.org/10.1111/j.1574-6976.1997.tb00340.x. doi: 10.1111/j.1574-6976.1997.tb00340.x
|
| [25] |
Rezza I, Salinas E, Calvente V, et al. (1997) Extraction of lithium from spodumene by bioleaching. Letters in applied microbiology 25: 172–176. https://doi.org/10.1046/j.1472-765x.1997.00199.x doi: 10.1046/j.1472-765x.1997.00199.x
|
| [26] |
Bosecker K (1997) Bioleaching: metal solubilization by microorganisms. FEMS Microbiology reviews 20: 591–604. https://doi.org/10.1111/j.1574-6976.1997.tb00340.x doi: 10.1111/j.1574-6976.1997.tb00340.x
|
| [27] | Brandl H (2001) Microbial leaching of metals. Biotechnology 10: 191–224. http://doi.wiley.com/10.1002/9783527620999.ch8k |
| [28] |
Bosecker K (2001) Microbial leaching in environmental clean-up programmes. Hydrometallurgy 59: 245–248. https://doi.org/10.1016/s0304-386x(00)00163-8 doi: 10.1016/s0304-386x(00)00163-8
|
| [29] |
Rojas-Chapana JA, Tributsch H (2001) Biochemistry of sulfur extraction in bio-corrosion of pyrite by Thiobacillus ferrooxidans. Hydrometallurgy 59: 291–300. https://doi.org/10.1016/S0304-386X(00)00185-7 doi: 10.1016/S0304-386X(00)00185-7
|
| [30] |
Sand W, Gehrke T, Jozsa PG, et al. (2001) Biochemistry of bacterial leaching—direct vs. indirect bioleaching. Hydrometallurgy 59: 159–175. https://doi.org/10.1016/s0304-386x(00)00180-8 doi: 10.1016/s0304-386x(00)00180-8
|
| [31] |
Johnson DB (2014) Biomining—biotechnologies for extracting and recovering metals from ores and waste materials. Current opinion in biotechnology 30: 24–31. https://doi.org/10.1016/j.copbio.2014.04.008 doi: 10.1016/j.copbio.2014.04.008
|
| [32] | Sala Cossich E, Granhen Tavares CR, Kakuta Ravagnani TM (2002) Biosorption of chromium (Ⅲ) by Sargassum sp. biomass. Electronic Journal of Biotechnology 5: 6–7. http://www.bioline.org.br/pdf?ej02024 |
| [33] |
Kumar KS, Dahms HU, Won EJ, et al. (2015) Microalgae–a promising tool for heavy metal remediation. Ecotoxicology and environmental safety 113: 329–352. https://doi.org/10.1016/j.ecoenv.2014.12.019. doi: 10.1016/j.ecoenv.2014.12.019
|
| [34] |
Zakhama S, Dhaouadi H, M'henni F (2011) Nonlinear modelisation of heavy metal removal from aqueous solution using Ulva lactuca algae. Bioresource technology 102: 786–796. https://doi.org/10.1016/j.biortech.2010.08.107 doi: 10.1016/j.biortech.2010.08.107
|
| [35] |
Balaji S, Kalaivani T, Shalini M, et al. (2016) Sorption sites of microalgae possess metal binding ability towards Cr (Ⅵ) from tannery effluents—a kinetic and characterization study. Desalination and Water Treatment 57: 14518–14529. https://doi.org/10.1080/19443994.2015.1064032 doi: 10.1080/19443994.2015.1064032
|
| [36] |
Jaafari J, Yaghmaeian K (2019) Optimization of heavy metal biosorption onto freshwater algae (Chlorella colonials) using response surface methodology (RSM). Chemosphere 217: 447–455. https://doi.org/10.1016/j.chemosphere.2018.10.205 doi: 10.1016/j.chemosphere.2018.10.205
|
| [37] |
Rahman Z, Singh VP (2019) The relative impact of toxic heavy metals (THMs) (arsenic (As), cadmium (Cd), chromium (Cr)(Ⅵ), mercury (Hg), and lead (Pb)) on the total environment: an overview. Environmental monitoring and assessment 191: 1–21. https://doi.org/10.1007/s10661-019-7528-7 doi: 10.1007/s10661-019-7528-7
|
| [38] |
Suvarapu LN, Baek SO (2017) Determination of heavy metals in the ambient atmosphere: A review. Toxicology and industrial health 33: 79–96. https://doi.org/10.1177/0748233716654827 doi: 10.1177/0748233716654827
|
| [39] |
Sun J, Cheng J, Yang Z, et al. (2015) Microstructures and functional groups of Nannochloropsis sp. cells with arsenic adsorption and lipid accumulation. Bioresource technology 194: 305–311. https://doi.org/10.1016/j.biortech.2015.07.041194:305–311 doi: 10.1016/j.biortech.2015.07.041194:305–311
|
| [40] |
Gómez-Jacinto V, García-Barrera T, Gómez-Ariza JL, et al. (2015) Elucidation of the defense mechanism in microalgae Chlorella sorokiniana under mercury exposure. Identification of Hg–phytochelatins. Chemico-Biological Interactions 238: 82–90. https://doi.org/10.1016/j.cbi.2015.06.013 doi: 10.1016/j.cbi.2015.06.013
|
| [41] |
Priatni S, Ratnaningrum D, Warya S, et al. (2018) Phycobiliproteins production and heavy metals reduction ability of Porphyridium sp. In IOP Conference Series: Earth and Environmental Science160: 012006. https://doi.org/10.1088/1755-1315/160/1/012006 doi: 10.1088/1755-1315/160/1/012006
|
| [42] |
Devars S, Avilés C, Cervantes C, et al. (2000) Mercury uptake and removal by Euglena gracilis. Archives of microbiology 174: 175–180. https://doi.org/10.1007/s002030000193 doi: 10.1007/s002030000193
|
| [43] |
Lu J, Li X, Hwang HY, et al. (2017) Coherent two-dimensional terahertz magnetic resonance spectroscopy of collective spin waves. Physical review letters 118: 207204. https://doi.org/10.1103/PhysRevLett.118.207204 doi: 10.1103/PhysRevLett.118.207204
|
| [44] |
Mathanker SK, Weckler PR, Bowser TJ (2013) X-ray applications in food and agriculture: a review. Transactions of the ASABE 56: 1227–1239. https://doi.org/10.13031/trans.56.9785 doi: 10.13031/trans.56.9785
|
| [45] |
Shao Y, Wang Y, Zhu D, et al. (2022) Measuring heavy metal ions in water using nature existed microalgae as medium based on terahertz technology. Journal of Hazardous Materials 435: 129028. https://doi.org/10.1016/j.jhazmat.2022.129028 doi: 10.1016/j.jhazmat.2022.129028
|
| [46] |
Krebs W, Brombacher C, Bosshard PP, et al. (1997) Microbial recovery of metals from solids. FEMS Microbiology reviews 20: 605–617. https://doi.org/10.1111/j.1574-6976.1997.tb00341.x doi: 10.1111/j.1574-6976.1997.tb00341.x
|
| [47] |
Pradhan N, Nathsarma KC, Rao KS, et al. (2008) Heap bioleaching of chalcopyrite: a review. Minerals Engineering 21: 355–365. https://doi.org/10.1016/j.mineng.2007.10.018 doi: 10.1016/j.mineng.2007.10.018
|
| [48] |
Ndlovu S (2008) Biohydrometallurgy for sustainable development in the African minerals industry. Hydrometallurgy 91: 20–27. https://doi.org/10.1016/j.hydromet.2007.11.007 doi: 10.1016/j.hydromet.2007.11.007
|
| [49] |
Rawlings DE (2004) Microbially-assisted dissolution of minerals and its use in the mining industry. Pure and Applied Chemistry 76: 847–859. https://doi.org/10.1351/pac200476040847 doi: 10.1351/pac200476040847
|
| [50] | Nkemnaso OC, Ejovwokoghene EF, Innocent CU (2018) Bio-mining: The Past, the Present and the Future. Bioscience and Bioengineering 4: 52–60. |
| [51] |
Hewedy MA, Rushdy AA, Kamal NM (2013) Bioleaching of rare earth elements and uranium from Sinai soil, Egypt using actinomycetes. The Egyptian Journal of Hospital Medicine 53: 909–917. https://doi.org/10.12816/0001653 doi: 10.12816/0001653
|
| [52] |
Hassanien W, Desouky O, Hussien S (2014) Bioleaching of some rare earth elements from Egyptian monazite using Aspergillus ficuum and Pseudomonas aeruginosa. Walailak journal of science and technology (WJST) 11: 809–823. https://doi.org/10.2004/WJST.V11I6.481 doi: 10.2004/WJST.V11I6.481
|
| [53] |
Al-Jabri H, Das P, Khan S, et al. (2020) Treatment of wastewaters by microalgae and the potential applications of the produced biomass—A review. Water 13: 27. https://doi.org/10.3390/w13010027 doi: 10.3390/w13010027
|
| [54] |
Abdel-Raouf N, Al-Homaidan AA, Ibraheem I (2012) Microalgae and wastewater treatment. Saudi journal of biological sciences 19: 257–275. https://doi.org/10.1016/j.sjbs.2012.04.005 doi: 10.1016/j.sjbs.2012.04.005
|
| [55] |
Rawat I, Ranjith Kumar R, Mutanda T, et al. (2011) Dual role of microalgae: Phytoremediation of domestic wastewater and biomass production for sustainable biofuels production. Applied Energy 88: 3411–3424. https://doi.org/10.1016/j.apenergy.2010.11.025 doi: 10.1016/j.apenergy.2010.11.025
|
| [56] |
El-Sheekh M, El-Dalatony MM, Thakur N, et al. (2022) Role of microalgae and cyanobacteria in wastewater treatment: Genetic engineering and omics approaches. International Journal of Environmental Science and Technology 19: 2173–2194. http://dx.doi.org/10.1007/s13762-021-03270-w doi: 10.1007/s13762-021-03270-w
|
| [57] |
Olguín EJ (2012) Dual purpose microalgae–bacteria-based systems that treat wastewater and produce biodiesel and chemical products within a Biorefinery. Biotechnology Advances 30: 1031–1046. https://doi.org/10.1016/j.biotechadv.2012.05.001 doi: 10.1016/j.biotechadv.2012.05.001
|
| [58] |
Fazal T, Mushtaq A, Rehman F, et al. (2018) Bioremediation of textile wastewater and successive biodiesel production using microalgae. Renewable and Sustainable Energy Reviews 82: 3107–3126. https://doi.org/10.1016/j.rser.2017.10.029 doi: 10.1016/j.rser.2017.10.029
|
| [59] |
Renuka N, Sood A, Prasanna R, et al. (2014) Phytoremediation of wastewaters: a synergistic approach using microalgae for bioremediation and biomass generation. International Journal of Environmental Science and Technology 12: 1443–1460. https://doi.org/10.1007/s13762-014-0700-2 doi: 10.1007/s13762-014-0700-2
|
| [60] |
Hussain F, Shah SZ, Ahmad H, et al. (2021) Microalgae an ecofriendly and sustainable wastewater treatment option: Biomass application in biofuel and bio-fertilizer production. A review. Renewable and Sustainable Energy Reviews 137: https://doi.org/10.1016/j.rser.2020.110603 doi: 10.1016/j.rser.2020.110603
|
| [61] |
Li X, Wu S, Yang C, et al. (2020) Microalgal and Duckweed Based Constructed Wetlands for Swine Wastewater Treatment: A Review. Bioresource Technology 310: https://doi.org/10.1016/j.biortech.2020.123858 doi: 10.1016/j.biortech.2020.123858
|
| [62] |
Ahmed S F, Mofijur M, Parisa T A, et al. (2022) Progress and challenges of contaminate removal from wastewater using microalgae biomass. Chemosphere 286: https://doi.org/10.1016/j.chemosphere.2021.131656 doi: 10.1016/j.chemosphere.2021.131656
|
| [63] |
Rawat I, Gupta SK, Shriwastav A, et al. (2016) Microalgae Applications in Wastewater Treatment. Green Energy and Technology 2016: 249–268. https://doi.org/10.1007/978-3-319-12334-9_13 doi: 10.1007/978-3-319-12334-9_13
|
| [64] |
Jones ER, Vliet MTH, Qadir M, et al (2021) Country level and gridded estimates of wastewater production, collection, treatment and reuse. Earth System Science Data 13: 237–254. https://doi.org/10.5194/essd-13-237-2021 doi: 10.5194/essd-13-237-2021
|
| [65] |
Zhou T, Li X, Zhang Q, et al. (2022) Ecotoxicological response of Spirulina platensis to coexisted copper and zinc in anaerobic digestion effluent. Science of the total environment 837: https://doi.org/10.1016/j.scitotenv.2022.155874 doi: 10.1016/j.scitotenv.2022.155874
|
| [66] |
Cheng H, Winayu BNR, Chu H, et al. (2020) Recent advancement on biological technologies and strategies for resource recovery from swine wastewater. Bioresource Technology 303: 122861. https://doi.org/10.1016/j.biortech.2020.122861 doi: 10.1016/j.biortech.2020.122861
|
| [67] |
Yu J, Hu H, Wu X, et al. (2020) Coupling of biochar-mediated absorption and algal-bacterial system to enhance nutrients recovery from swine wastewater. Science of The Total Environment 701: 134935. https://doi.org/10.1016/j.scitotenv.2019.134935 doi: 10.1016/j.scitotenv.2019.134935
|
| [68] |
Nguyen HT, Yoon Y, Ngo HH, et al. (2021) The application of microalgae in removing organic micropollutants in wastewater. Critical Reviews in Environmental Science and Technology 51: 1187-1220. https://doi.org/10.1080/10643389.2020.1753633 doi: 10.1080/10643389.2020.1753633
|
| [69] |
Cao Y, Shao P, Chen Y, et al. (2021) A critical review of the recovery of rare earth elements from wastewater by algae for resources recycling technologies. Resources, Conservation and Recycling 169: 105519. https://doi.org/10.1016/j.resconrec.2021.105519 doi: 10.1016/j.resconrec.2021.105519
|
| [70] |
Moran, S (2018) An Applied Guide to Water and Effluent Treatment Plant Design. Butterworth-Heinemann. https://doi.org/10.1016/B978-0-12-811309-7.00020-5 doi: 10.1016/B978-0-12-811309-7.00020-5
|
| [71] |
Jais NM, Mohamed RMSR, Al-Gheethi AA, et al. (2017) The dual roles of phytoremediation of wet market wastewater for nutrients and heavy metals removal and microalgae biomass production. Clean Technologies and Environmental Policy 19: 37–52. https://doi.org/10.1007/s10098-016-1235-7 doi: 10.1007/s10098-016-1235-7
|
| [72] |
Wang Y, Ho SH, Cheng CL, et al. (2016) Perspectives on the feasibility of using microalgae for industrial wastewater treatment. Bioresource Technology 222: 485–497. https://doi.org/10.1016/j.biortech.2016.09.106 doi: 10.1016/j.biortech.2016.09.106
|
| [73] |
Chai WS, Tan WG, Munawaroh HSH, et al. (2021) Multifaceted roles of microalgae in the application of wastewater biotreatment: a review. Environmental Pollution 269: 116236. https://doi.org/10.1016/j.envpol.2020.116236 doi: 10.1016/j.envpol.2020.116236
|
| [74] |
Shahid A, Malik S, Zhu H, et al. (2020) Cultivating microalgae in wastewater for biomass production, pollutant removal, and atmospheric carbon mitigation; a review. Science of the Total Environment 704: 135303. https://doi.org/10.1016/j.scitotenv.2019.135303 doi: 10.1016/j.scitotenv.2019.135303
|
| [75] |
Gao H, Scherson YD, Wells GF (2014) Towards energy neutral wastewater treatment: methodology and state of the art. Environmental science: Processes & impacts 16: 1223–1246. https://doi.org/10.1039/C4EM00069B doi: 10.1039/C4EM00069B
|
| [76] |
De Corte S, Hennebel T, De Gusseme B, et al. (2012) Bio‐palladium: from metal recovery to catalytic applications. Microbial Biotechnology 5: 5–17. https://doi.org/10.1111/j.1751-7915.2011.00265.x doi: 10.1111/j.1751-7915.2011.00265.x
|
| [77] |
Wang L, Xiao H, He N, et al. (2019) Biosorption and Biodegradation of the Environmental Hormone Nonylphenol by Four Marine Microalgae. Scientific reports 9: 5277. https://doi.org/10.1038/s41598-019-41808-8 doi: 10.1038/s41598-019-41808-8
|
| [78] |
Della Greca M, Pinto G, Pistillo P, et al. (2008) Biotransformation of ethinylestradiol by microalgae. Chemosphere 70: 2047–2053. https://doi.org/10.1016/j.chemosphere.2007.09.011 doi: 10.1016/j.chemosphere.2007.09.011
|
| [79] |
Kržanović D, Conić V, Bugarin D, et al. (2019) Maximizing economic performance in the mining industry by applying bioleaching technology for extraction of polymetallic mineral deposits. Minerals 9: 400. https://doi.org/10.3390/min9070400 doi: 10.3390/min9070400
|
| [80] |
Kumar A, Dimitrakopoulos R (2019) Application of simultaneous stochastic optimization with geometallurgical decisions at a copper–gold mining complex. Mining Technology 128: 88–105. https://doi.org/10.1080/25726668.2019.1575053 doi: 10.1080/25726668.2019.1575053
|
| [81] |
Mulligan CN, Kamali M, Gibbs BF (2004) Bioleaching of heavy metals from a low-grade mining ore using Aspergillus niger. Journal of Hazardous Materials 110: 77–84. https://doi.org/10.1016/j.jhazmat.2004.02.040 doi: 10.1016/j.jhazmat.2004.02.040
|
| [82] |
di Cicco MR, Iovinella M, Palmieri, et al. (2021) Extremophilic microalgae Galdieria gen. for urban wastewater treatment: current state, the case of "POWER" system, and prospects. Plants 10: 2343. https://doi.org/10.3390/plants10112343 doi: 10.3390/plants10112343
|
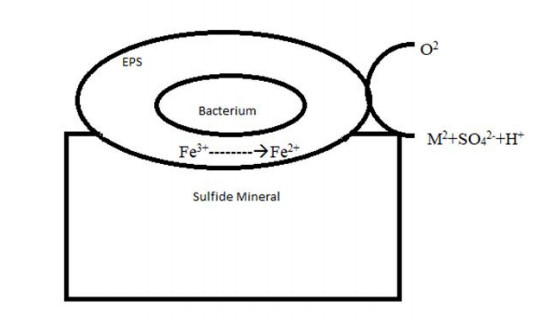
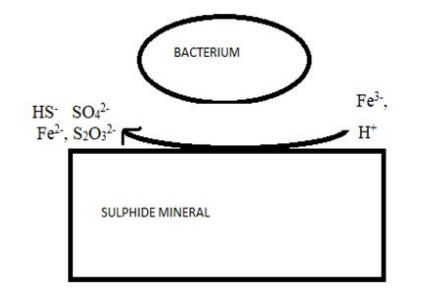
Figures(6) / Tables(2)

Susmita Mukherjee, Sharanya Paul, Shreya Bhattacharjee, Somava Nath, Upasana Sharma, Sonali Paul. Bioleaching of critical metals using microalgae[J]. AIMS Environmental Science, 2023, 10(2): 226-244. doi: 10.3934/environsci.2023013







 DownLoad:
DownLoad: