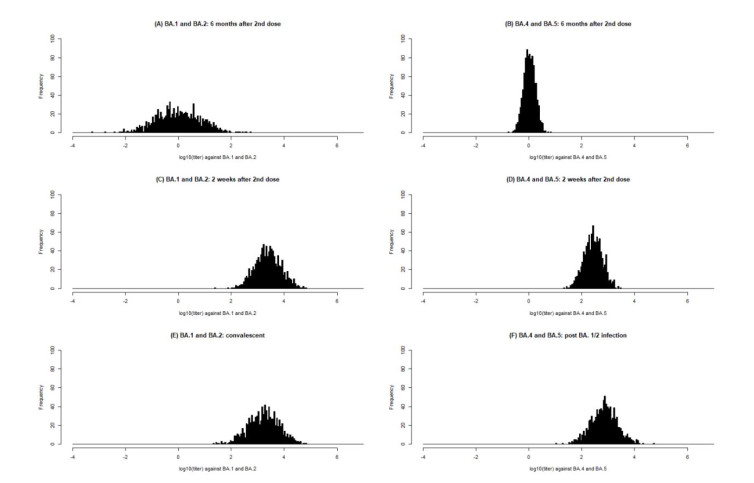
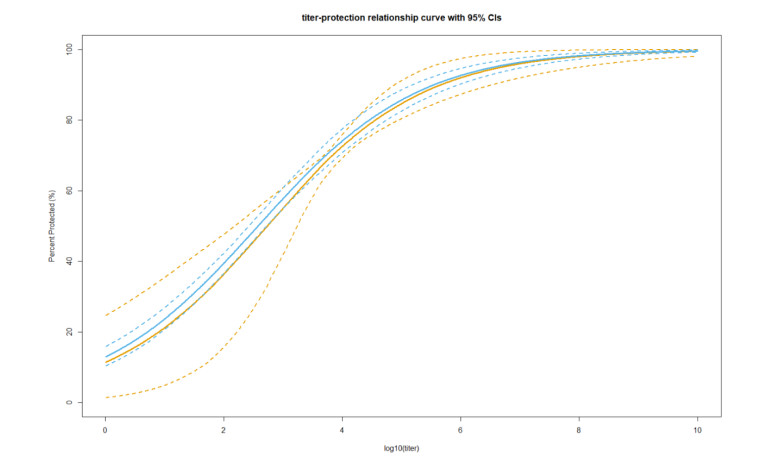
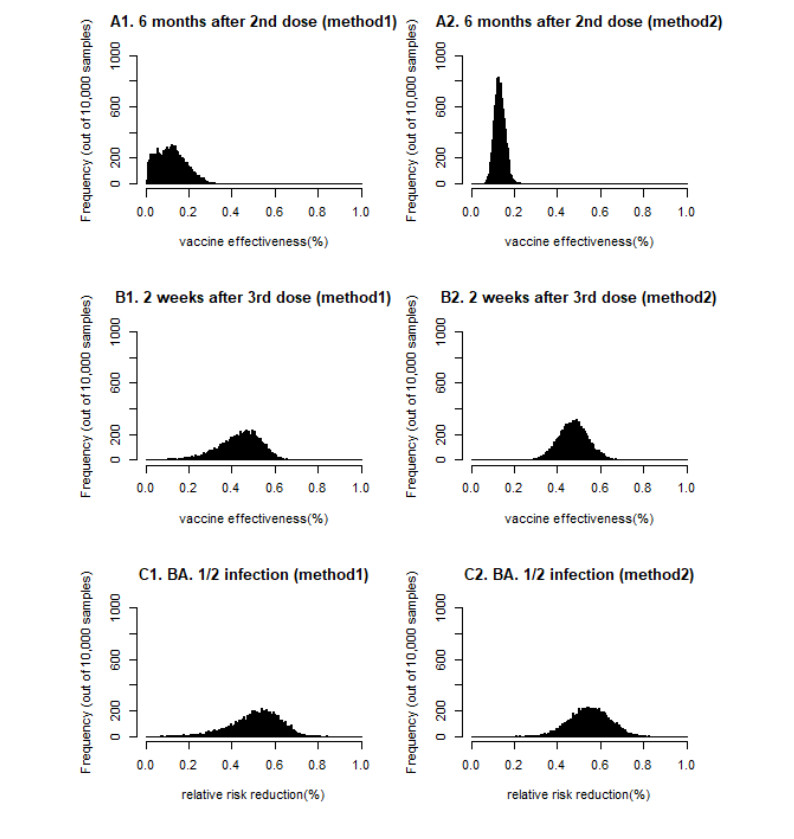
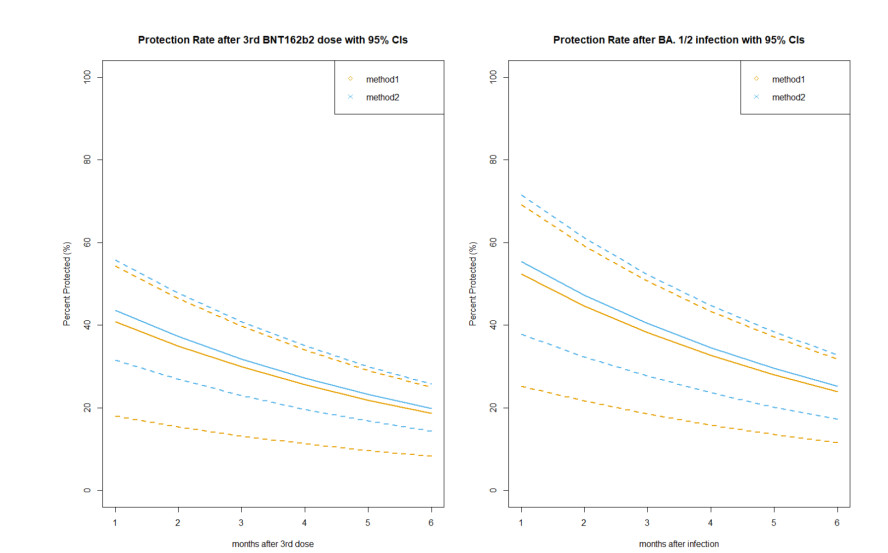
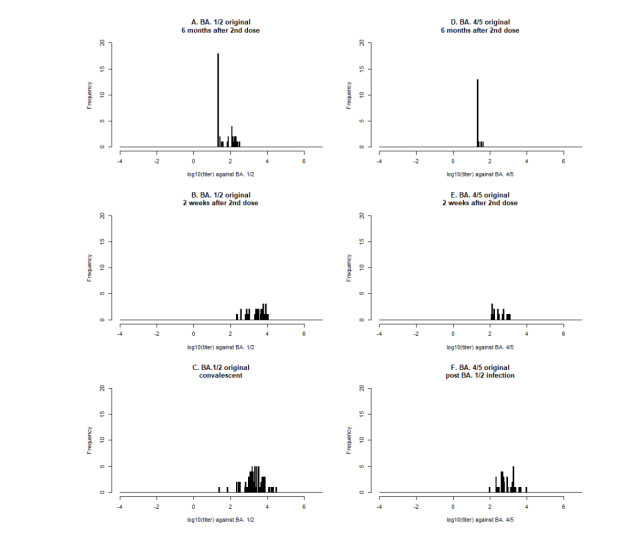
With continuing emergence of new SARS-CoV-2 variants, understanding the proportion of the population protected against infection is crucial for public health risk assessment and decision-making and so that the general public can take preventive measures. We aimed to estimate the protection against symptomatic illness caused by SARS-CoV-2 Omicron variants BA.4 and BA.5 elicited by vaccination against and natural infection with other SARS-CoV-2 Omicron subvariants. We used a logistic model to define the protection rate against symptomatic infection caused by BA.1 and BA.2 as a function of neutralizing antibody titer values. Applying the quantified relationships to BA.4 and BA.5 using two different methods, the estimated protection rate against BA.4 and BA.5 was 11.3% (95% confidence interval [CI]: 0.01–25.4) (method 1) and 12.9% (95% CI: 8.8–18.0) (method 2) at 6 months after a second dose of BNT162b2 vaccine, 44.3% (95% CI: 20.0–59.3) (method 1) and 47.3% (95% CI: 34.1–60.6) (method 2) at 2 weeks after a third BNT162b2 dose, and 52.3% (95% CI: 25.1–69.2) (method 1) and 54.9% (95% CI: 37.6–71.4) (method 2) during the convalescent phase after infection with BA.1 and BA.2, respectively. Our study indicates that the protection rate against BA.4 and BA.5 are significantly lower compared with those against previous variants and may lead to substantial morbidity, and overall estimates were consistent with empirical reports. Our simple yet practical models enable prompt assessment of public health impacts posed by new SARS-CoV-2 variants using small sample-size neutralization titer data to support public health decisions in urgent situations.
Citation: Yuta Okada, Taishi Kayano, Asami Anzai, Tong Zhang, Hiroshi Nishiura. Protection against SARS-CoV-2 BA.4 and BA.5 subvariants via vaccination and natural infection: A modeling study[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 2530-2543. doi: 10.3934/mbe.2023118
With continuing emergence of new SARS-CoV-2 variants, understanding the proportion of the population protected against infection is crucial for public health risk assessment and decision-making and so that the general public can take preventive measures. We aimed to estimate the protection against symptomatic illness caused by SARS-CoV-2 Omicron variants BA.4 and BA.5 elicited by vaccination against and natural infection with other SARS-CoV-2 Omicron subvariants. We used a logistic model to define the protection rate against symptomatic infection caused by BA.1 and BA.2 as a function of neutralizing antibody titer values. Applying the quantified relationships to BA.4 and BA.5 using two different methods, the estimated protection rate against BA.4 and BA.5 was 11.3% (95% confidence interval [CI]: 0.01–25.4) (method 1) and 12.9% (95% CI: 8.8–18.0) (method 2) at 6 months after a second dose of BNT162b2 vaccine, 44.3% (95% CI: 20.0–59.3) (method 1) and 47.3% (95% CI: 34.1–60.6) (method 2) at 2 weeks after a third BNT162b2 dose, and 52.3% (95% CI: 25.1–69.2) (method 1) and 54.9% (95% CI: 37.6–71.4) (method 2) during the convalescent phase after infection with BA.1 and BA.2, respectively. Our study indicates that the protection rate against BA.4 and BA.5 are significantly lower compared with those against previous variants and may lead to substantial morbidity, and overall estimates were consistent with empirical reports. Our simple yet practical models enable prompt assessment of public health impacts posed by new SARS-CoV-2 variants using small sample-size neutralization titer data to support public health decisions in urgent situations.

| [1] |
N. Zhu, D. Zhang, W. Wang, X. Li, B. Yang, J. Song, et al., A novel coronavirus from patients with pneumonia in China, 2019, New Engl. J. Med., 382 (2020), 727–733. https://doi.org/10.1056/NEJMoa2001017 doi: 10.1056/NEJMoa2001017

|
| [2] |
E. A. Meyerowitz, A. Richterman, R. T. Gandhi, P. E. Sax, Transmission of SARS-CoV-2: A review of viral, host, and environmental factors, 174 (2021), 69–79. https://doi.org/10.7326/M20-5008 doi: 10.7326/M20-5008
|
| [3] |
A. Núñez-Delgado, E. Bontempi, M. Coccia, M. Kumar, K. Farkas, J. L. Domingo, SARS-CoV-2 and other pathogenic microorganisms in the environment, Environ. Res., 201 (2021), 111606. https://doi.org/10.1016/j.envres.2021.111606 doi: 10.1016/j.envres.2021.111606
|
| [4] |
M. Cevik, J. L. Marcus, C. Buckee, T. C.Smith, Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) transmission dynamics should inform policy, Clin. Infect. Dis., 73 (2021), S170–S176. https://doi.org/10.1093/cid/ciaa1442 doi: 10.1093/cid/ciaa1442
|
| [5] |
D. Nash, M. S. Rane, M. M. Robertson, M. Chang, S. G. Kulkarni, R. Zimba, et al., SARS-CoV-2 incidence and risk factors in a national, community-based prospective cohort of U.S. adults, Clin. Infect. Dis., (2022). https://doi.org/10.1101/2021.02.12.21251659 doi: 10.1101/2021.02.12.21251659
|
| [6] |
M. Coccia, Factors determining the diffusion of COVID-19 and suggested strategy to prevent future accelerated viral infectivity similar to COVID, Sci. Total. Environ., 729 (2020), 138474. https://doi.org/10.1016/j.scitotenv.2020.138474 doi: 10.1016/j.scitotenv.2020.138474
|
| [7] |
M. Coccia, COVID-19 pandemic over 2020 (with lockdowns) and 2021 (with vaccinations): similar effects for seasonality and environmental factors, Environ. Res., 208 (2022), 112711. https://doi.org/10.1016/j.envres.2022.112711 doi: 10.1016/j.envres.2022.112711
|
| [8] |
M. Coccia, Optimal levels of vaccination to reduce COVID-19 infected individuals and deaths: A global analysis, Environ. Res., 204 (2022). https://doi.org/10.1016/j.envres.2021.112314 doi: 10.1016/j.envres.2021.112314
|
| [9] |
M. Coccia, Preparedness of countries to face COVID-19 pandemic crisis: Strategic positioning and factors supporting effective strategies of prevention of pandemic threats, Environ. Res., 203 (2022), 111678. https://doi.org/10.1016/j.envres.2021.111678 doi: 10.1016/j.envres.2021.111678
|
| [10] |
I. Benati, M. Coccia, Global analysis of timely COVID-19 vaccinations: Improving governance to reinforce response policies for pandemic crises, Int. J. Health Gov., 27 (2022), 240–253. https://doi.org/10.1108/IJHG-07-2021-0072 doi: 10.1108/IJHG-07-2021-0072
|
| [11] |
L. Matrajt, J. Eaton, T. Leung, E. R. Brown, Vaccine optimization for COVID-19: Who to vaccinate first?, Sci. Adv., 7 (2021). https://doi.org/10.1126/sciadv.abf1374 doi: 10.1126/sciadv.abf1374
|
| [12] |
L. Thul, W. Powell, Stochastic optimization for vaccine and testing kit allocation for the COVID-19 pandemic, Eur. J. Oper. Res., 304 (2023), 325–338. https://doi.org/10.1016/j.ejor.2021.11.007 doi: 10.1016/j.ejor.2021.11.007
|
| [13] |
W. Cao, J. Zhu, X. Wang, X. Tong, Y. Tian, H. Dai, et al., Optimizing spatio-temporal allocation of the COVID-19 vaccine under different epidemiological landscapes, Front. Public Health, 10 (2022), 1828. https://doi.org/10.3389/fpubh.2022.921855 doi: 10.3389/fpubh.2022.921855
|
| [14] |
M. Bicher, C. Rippinger, M. Zechmeister, B. Jahn, G. Sroczynski, N. Mühlberger, et al., An iterative algorithm for optimizing COVID-19 vaccination strategies considering unknown supply, PLoS One, 17 (2022), e0265957. https://doi.org/10.1371/journal.pone.0265957 doi: 10.1371/journal.pone.0265957
|
| [15] |
L. Matrajt, J. Eaton, T. Leung, D. Dimitrov, J. T. Schiffer, D. A. Swan, et al., Optimizing vaccine allocation for COVID-19 vaccines shows the potential role of single-dose vaccination, Nat. Commun., 12 (2021), 1–18. https://doi.org/10.1038/s41467-021-23761-1 doi: 10.1038/s41467-021-23761-1
|
| [16] | M. Coccia, Comparative critical decisions in management, in Global Encyclopedia of Public Administration, Public Policy, and Governance, (2020), 1–10. https://doi.org/10.1007/978-3-319-31816-5_3969-1 |
| [17] |
M. Coccia, Improving preparedness for next pandemics: Max level of COVID-19 vaccinations without social impositions to design effective health policy and avoid flawed democracies, Environ. Res., 213 (2022), 113566. https://doi.org/10.1016/j.envres.2022.113566 doi: 10.1016/j.envres.2022.113566
|
| [18] |
M. Coccia, Pandemic prevention: Lessons from COVID-19, Encyclopedia, 1 (2021), 433–444. https://doi.org/10.3390/encyclopedia1020036 doi: 10.3390/encyclopedia1020036
|
| [19] |
R. P. Kumar, P. K. Santra, G. S. Mahapatra, Global stability and analysing the sensitivity of parameters of a multiple-susceptible population model of SARS-CoV-2 emphasising vaccination drive, Math. Comput. Simul., 203 (2023), 741–766. https://doi.org/10.1016/j.matcom.2022.07.012 doi: 10.1016/j.matcom.2022.07.012
|
| [20] |
Z. Wang, F. Schmidt, Y. Weisblum, F. Muecksch, C. O. Barnes, S. Finkin, et al., mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants, Nature, 592 (2021), 616–622. https://doi.org/10.1038/s41586-021-03324-6 doi: 10.1038/s41586-021-03324-6
|
| [21] |
Y. Liu, J. Liu, H. Xia, X. Zhang, C. R. Fontes-Garfias, K. A. Swanson, et al., Neutralizing activity of BNT162b2-Elicited serum, N. Engl. J. Med., 384 (2021), 1466–1468. https://doi.org/10.1056/NEJMc2102017 doi: 10.1056/NEJMc2102017
|
| [22] |
R. E. Chen, X. Zhang, J. B. Case, E. S. Winkler, Y. Liu, L. A. VanBlargan, et al., Resistance of SARS-CoV-2 variants to neutralization by monoclonal and serum-derived polyclonal antibodies, Nat. Med., 27 (2021), 717–726.https://doi.org/10.1038/s41591-021-01294-w doi: 10.1038/s41591-021-01294-w
|
| [23] |
P. Wang, M. S. Nair, L. Liu, S. Iketani, Y. Luo, Y. Guo, et al., Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7, Nature, 593 (2021), 130–135. https://doi.org/10.1038/s41586-021-03398-2 doi: 10.1038/s41586-021-03398-2
|
| [24] |
D. S. Khoury, D. Cromer, A. Reynaldi, T. E. Schlub, A. K. Wheatley, J. A. Juno, et al., Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection, Nat. Med., 27 (2021), 1205–1211. https://doi.org/10.1038/s41591-021-01377-8 doi: 10.1038/s41591-021-01377-8
|
| [25] |
L. Wang, M. H. Kainulainen, N. Jiang, H. Di, G. Bonenfant, L. Mills, et al., Differential neutralization and inhibition of SARS-CoV-2 variants by antibodies elicited by COVID-19 mRNA vaccines, Nat. Commun., 13 (2022), 1–10. https://doi.org/10.1038/s41467-022-31929-6 doi: 10.1038/s41467-022-31929-6
|
| [26] |
S. P. Andeweg, B. de Gier, D. Eggink, C. van den Ende, N. van Maarseveen, L. Ali, et al., Protection of COVID-19 vaccination and previous infection against Omicron BA.1, BA.2 and Delta SARS-CoV-2 infections, Nat. Commun., 13 (2022), 1–9. https://doi.org/10.1038/s41467-022-31838-8 doi: 10.1038/s41467-022-31838-8
|
| [27] |
N. Andrews, J. Stowe, F. Kirsebom, S. Toffa, T. Rickeard, E. Gallagher, et al., Covid-19 Vaccine effectiveness against the omicron (B.1.1.529) variant, N. Engl. J. Med., 386 (2022), 1532–1546. https://doi.org/10.1056/NEJMoa2119451 doi: 10.1056/NEJMoa2119451
|
| [28] |
G. Regev-Yochay, T. Gonen, M. Gilboa, M. Mandelboim, V. Indenbaum, S. Amit, et al., Efficacy of a fourth dose of Covid-19 mRNA vaccine against Omicron, N. Engl. J. Med., 386 (2022), 1377–1380. https://doi.org/10.1056/NEJMc2202542 doi: 10.1056/NEJMc2202542
|
| [29] |
E. Nanishi, M. E. McGrath, T. R. O'Meara, S. Barman, J. Yu, H. Wan, et al., mRNA booster vaccination protects aged mice against the SARS-CoV-2 Omicron variant, Commun. Biol., 5 (2022), 1–8. https://doi.org/10.1038/s42003-022-03765-3 doi: 10.1038/s42003-022-03765-3
|
| [30] |
N. P. Hachmann, J. Miller, A. Y. Collier, J. D. Ventura, J. Yu, M. Rowe, et al., Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5, N. Engl. J. Med., 387 (2022). https://doi.org/10.1101/2022.05.16.22275151 doi: 10.1101/2022.05.16.22275151
|
| [31] |
Y. Cao, A. Yisimayi, F. Jian, W. Song, T. Xiao, L. Wang, et al., BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection, Nature, 608 (2022), 1–3. https://doi.org/10.1038/s41586-022-04980-y doi: 10.1038/s41586-022-04980-y
|
| [32] |
Q. Wang, Y. Guo, S. Iketani, M. S. Nair, Z. Li, H. Mohri, et al., Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4, & BA.5, Nature, 608 (2022), 1–3. https://doi.org/10.1038/s41586-022-05053-w doi: 10.1038/s41586-022-05053-w
|
| [33] |
M. Huang, L. Wu, A. Zheng, Y. Xie, Q. He, X. Rong, et al., Atlas of currently-available human neutralizing antibodies against SARS-CoV-2 and escape by Omicron sub-variants BA.1/BA.1.1/BA.2/BA.3, Immunity, 55 (2022). https://doi.org/10.1016/j.immuni.2022.06.005 doi: 10.1016/j.immuni.2022.06.005
|
| [34] |
K. Khan, F. Karim, Y. Ganga, M. Bernstein, Z. Jule, K. Reedoy, et al., Omicron BA.4/BA.5 escape neutralizing immunity elicited by BA.1 infection, Nat. Commun., 13 (2022), 1–7. https://doi.org/10.1038/s41467-022-32396-9 doi: 10.1038/s41467-022-32396-9
|
| [35] |
L. Bellusci, G. Grubbs, F. T. Zahra, D. Forgacs, H. Golding, T. M. Ross, et al., Antibody affinity and cross-variant neutralization of SARS-CoV-2 Omicron BA.1, BA.2 and BA.3 following third mRNA vaccination, Nat. Commun., 13 (2022), 1–9. https://doi.org/10.1038/s41467-022-32298-w doi: 10.1038/s41467-022-32298-w
|
| [36] |
L. Qin, P. B. Gilbert, L. Corey, M. J. McElrath, S. G. Self, A framework for assessing immunological correlates of protection in vaccine trials, J. Infect. Dis., 196 (2007), 1304–1312. https://doi.org/10.1086/522428 doi: 10.1086/522428
|
| [37] |
J. Storsaeter, H. O. Hallander, L. Gustafsson, P. Olin, Levels of anti-pertussis antibodies related to protection after household exposure to Bordetella pertussis, Vaccine, 16 (1998), 1907–1916. https://doi.org/10.1016/S0264-410X(98)00227-8 doi: 10.1016/S0264-410X(98)00227-8
|
| [38] |
Y J. Yu, A. Y. Collier, M. Rowe, F. Mardas, J. D. Ventura, H. Wan, et al., Neutralization of the SARS-CoV-2 Omicron BA.1 and BA.2 Variants, N. Engl. J. Med., 386 (2022), 1579–1580. https://doi.org/10.1056/NEJMc2201849 doi: 10.1056/NEJMc2201849
|
| [39] |
S. de Montigny, B. J. S. Adamson, B. R. Mâsse, L. P. Garrison, J. G. Kublin, P. B. Gilbert, et al., Projected effectiveness and added value of HIV vaccination campaigns in South Africa: A modeling study, Sci. Rep., 8 (2018), 1–12. https://doi.org/10.1038/s41598-018-24268-4 doi: 10.1038/s41598-018-24268-4
|
| [40] |
K. M. Andersson, J. Stover, The potential impact of a moderately effective HIV vaccine with rapidly waning protection in South Africa and Thailand, Vaccine, 29 (2011), 6092–6099. https://doi.org/10.1016/j.vaccine.2011.06.060 doi: 10.1016/j.vaccine.2011.06.060
|
| [41] |
J. A. C. Hontelez, N. Nagelkerke, T. Bärnighausen, R. Bakker, F. Tanser, M. L. Newell, et al., The potential impact of RV144-like vaccines in rural South Africa: A study using the STDSIM microsimulation model, Vaccine, 29 (2011), 6100–6106. https://doi.org/10.1016/j.vaccine.2011.06.059 doi: 10.1016/j.vaccine.2011.06.059
|
| [42] |
L. B. Shrestha, C. Foster, W. Rawlinson, N. Tedla, R. A. Bull, C. A. R. Bull, Evolution of the SARS-CoV-2 omicron variants BA.1 to BA.5: Implications for immune escape and transmission, Rev. Med. Virol., 32 (2022), e2381. https://doi.org/10.1002/rmv.2381 doi: 10.1002/rmv.2381
|
| [43] |
A. Tuekprakhon, J. Huo, R. Nutalai, A. Dijokaite-Guraliuc, D. Zhou, H. M. Ginn, et al., Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum, Cell, 185 (2022), 2422–2433. https://doi.org/10.1016/j.cell.2022.06.005 doi: 10.1016/j.cell.2022.06.005
|
| [44] | H. Kared, A. S. Wolf, A. Alirezaylavasani, A. Ravussin, G. Solum, T. T. Tran, et al., Immune responses in Omicron SARS-CoV-2 breakthrough infection in vaccinated adults, Nat. Commun., 13 (2022), 1–12. |
| [45] |
H. N. Altarawneh, H. Chemaitelly, H. H. Ayoub, M. R. Hasan, P. Coyle, H. M. Yassine, et al., Protection of SARS-CoV-2 natural infection against reinfection with the Omicron BA.4 or BA.5 subvariants, MedRxiv, (2022). https://doi.org/10.1101/2022.07.11.22277448 doi: 10.1101/2022.07.11.22277448
|
| [46] |
J. Malato, R. M. Ribeiro, P. P. Leite, P. Casaca, E. Fernandes, C. Antunes, et al., Risk of BA.5 infection in individuals exposed to prior SARS-CoV-2 variants, MedRxiv, (2022). https://doi.org/10.1101/2022.07.27.22277602 doi: 10.1101/2022.07.27.22277602
|
| [47] |
C. H. Hansen, N. U. Friis, P. Bager, M. Stegger, J. Fonager, A. Fomsgaard, et al., Risk of reinfection, vaccine protection, and severity of infection with the BA.5 Omicron subvariant: A danish nation-wide population-based study, SSRN Electron. J., (2022). https://doi.org/10.2139/ssrn.4165630 doi: 10.2139/ssrn.4165630
|
| [48] |
J. T. Castro, P. Azevedo, M. J. Fumagalli, N. S. Hojo-Souza, N. Salazar, G. G. Almeida, et al., Promotion of neutralizing antibody-independent immunity to wild-type and SARS-CoV-2 variants of concern using an RBD-Nucleocapsid fusion protein, Nat. Commun., 13 (2022), 1–16. https://doi.org/10.1038/s41467-022-32547-y doi: 10.1038/s41467-022-32547-y
|
| [49] |
M. Reinscheid, H. Luxenburger, V. Karl, A. Graeser, S. Giese, K. Ciminski, et al., COVID-19 mRNA booster vaccine induces transient CD8+ T effector cell responses while conserving the memory pool for subsequent reactivation, Nat. Commun., 13 (2022), 1–11. https://doi.org/10.1038/s41467-022-32324-x doi: 10.1038/s41467-022-32324-x
|
Figures(5)

Yuta Okada, Taishi Kayano, Asami Anzai, Tong Zhang, Hiroshi Nishiura. Protection against SARS-CoV-2 BA.4 and BA.5 subvariants via vaccination and natural infection: A modeling study[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 2530-2543. doi: 10.3934/mbe.2023118







 DownLoad:
DownLoad: