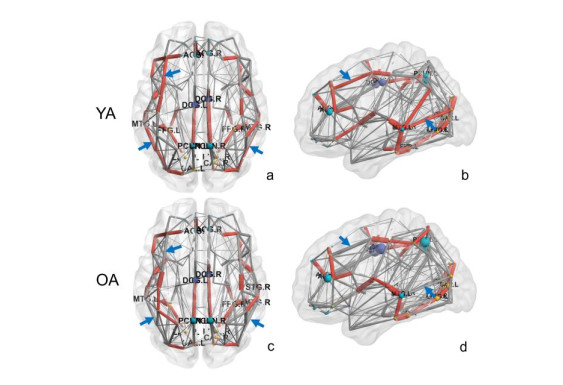
The approach of graph-based diffusion tensor imaging (DTI) networks has been used to explore the complicated structural connectivity of brain aging. In this study, the changes of DTI networks of brain aging were quantitatively and qualitatively investigated by comparing the characteristics of brain network. A cohort of 60 volunteers was enrolled and equally divided into young adults (YA) and older adults (OA) groups. The network characteristics of critical nodes, path length (Lp), clustering coefficient (Cp), global efficiency (Eglobal), local efficiency (Elocal), strength (Sp), and small world attribute (σ) were employed to evaluate the DTI networks at the levels of whole brain, bilateral hemispheres and critical brain regions. The correlations between each network characteristic and age were predicted, respectively. Our findings suggested that the DTI networks produced significant changes in network configurations at the critical nodes and node edges for the YA and OA groups. The analysis of whole brains network revealed that Lp, Cp increased (p < 0.05, positive correlation), Eglobal, Elocal, Sp decreased (p < 0.05, negative correlation), and σ unchanged (p ≥ 0.05, non-correlation) between the YA and OA groups. The analyses of bilateral hemispheres and brain regions showed similar results as that of the whole-brain analysis. Therefore the proposed scheme of DTI networks could be used to evaluate the WM changes of brain aging, and the network characteristics of critical nodes exhibited valuable indications for WM degeneration.
Citation: Song Xu, Xufeng Yao, Liting Han, Yuting Lv, Xixi Bu, Gan Huang, Yifeng Fan, Tonggang Yu, Gang Huang. Brain network analyses of diffusion tensor imaging for brain aging[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6066-6078. doi: 10.3934/mbe.2021303
The approach of graph-based diffusion tensor imaging (DTI) networks has been used to explore the complicated structural connectivity of brain aging. In this study, the changes of DTI networks of brain aging were quantitatively and qualitatively investigated by comparing the characteristics of brain network. A cohort of 60 volunteers was enrolled and equally divided into young adults (YA) and older adults (OA) groups. The network characteristics of critical nodes, path length (Lp), clustering coefficient (Cp), global efficiency (Eglobal), local efficiency (Elocal), strength (Sp), and small world attribute (σ) were employed to evaluate the DTI networks at the levels of whole brain, bilateral hemispheres and critical brain regions. The correlations between each network characteristic and age were predicted, respectively. Our findings suggested that the DTI networks produced significant changes in network configurations at the critical nodes and node edges for the YA and OA groups. The analysis of whole brains network revealed that Lp, Cp increased (p < 0.05, positive correlation), Eglobal, Elocal, Sp decreased (p < 0.05, negative correlation), and σ unchanged (p ≥ 0.05, non-correlation) between the YA and OA groups. The analyses of bilateral hemispheres and brain regions showed similar results as that of the whole-brain analysis. Therefore the proposed scheme of DTI networks could be used to evaluate the WM changes of brain aging, and the network characteristics of critical nodes exhibited valuable indications for WM degeneration.

| [1] | O. Carmichael, S. Lockhart, The role of diffusion tensor imaging in the study of cognitive aging, Curr. Top. Behav. Neurosci., 11 (2012), 289-320. |
| [2] |
A. Qiu, S. Mori, M. I. Miller, Diffusion tensor imaging for understanding brain development in early life, Annu. Rev. Psychol., 66 (2015), 853-876. doi: 10.1146/annurev-psych-010814-015340

|
| [3] |
S. Uda, M. Matsui, C. Tanaka, A. Uematsu, K. Miura, I. Kawana, et al., Normal development of human brain white matter from infancy to early adulthood: a diffusion tensor imaging study, Dev. Neurosci., 37 (2015), 182-194. doi: 10.1159/000373885
|
| [4] |
S. Pujol, W. Wells, C. Pierpaoli, C. Brun, J. Gee, G. Cheng, et al., The DTI challenge: toward standardized evaluation of diffusion tensor imaging tractography for neurosurgery, J. Neuroimaging, 25 (2015), 875-882. doi: 10.1111/jon.12283
|
| [5] | B. D. Le, J. F. Mangin, C. Poupon, C. A. Clark, S. Pappata, N, Molko, et al., Diffusion tensor imaging: concepts and applications, J. Magn. Reson. Imaging: Off. J. Int. Society Magn. Reson. Med., 13 (2001), 534-546. |
| [6] |
R. Xue, P. C. M. van Zijl, B. J. Crain, M. Solaiyappan, S. Mori, In vivo three-dimensional reconstruction of rat brain axonal projections by diffusion tensor imaging, Magn. Reson. Med.: Off. J. Int. Soc. Magn. Reson. Med., 42 (1999), 1123-1127. doi: 10.1002/(SICI)1522-2594(199912)42:6<1123::AID-MRM17>3.0.CO;2-H
|
| [7] | L. Bonilha, E. Gleichgerrcht, J. Fridriksson, C. Rorden, J. L. Breedlove, T. Nesland, et al., Reproducibility of the structural brain connectome derived from diffusion tensor imaging, PLoS One, 10 (2015), e0135247. |
| [8] |
A. L. Alexander, J. E. Lee, M. Lazar, A. S. Field, Diffusion tensor imaging of the brain, Neurotherapeutics, 4 (2007), 316-329. doi: 10.1016/j.nurt.2007.05.011
|
| [9] |
P. C. Sundgren, Q. Dong, D. Gomez-Hassan, S. K. Mukherji, P. Maly, R. Welsh, Diffusion tensor imaging of the brain: review of clinical applications, Neuroradiology, 46 (2004), 339-350. doi: 10.1007/s00234-003-1114-x
|
| [10] |
C. Yan, G. Gong, J. Wang, D. Wang, D. Liu, C. Zhu, et al., Sex-and brain size-related small-world structural cortical networks in young adults: a DTI tractography study, Cereb. Cortex, 21 (2011), 449-458. doi: 10.1093/cercor/bhq111
|
| [11] |
S. H. Jang, S. H. Cho, M. C. Chang, Age-related degeneration of the fornix in the human brain: a diffusion tensor imaging study, Int. J. Neurosci., 121 (2011), 94-100. doi: 10.3109/00207454.2010.531894
|
| [12] |
S. H. Jang, Y. H. Kwon, M. Y. Lee, J. R. Kim, J. P. Seo, Aging of the cingulum in the human brain: Preliminary study of a diffusion tensor imaging study, Neurosci. Lett., 610 (2016), 213-217. doi: 10.1016/j.neulet.2015.11.018
|
| [13] |
O. Abe, H. Yamasue, S. Aoki, M. Suga, H. Yamada, K. Kasai, et al., Aging in the CNS: comparison of gray/white matter volume and diffusion tensor data, Neurobiol. Aging, 29 (2008), 102-116. doi: 10.1016/j.neurobiolaging.2006.09.003
|
| [14] | H. Liu, L. Wang, Z. Geng, Q. Zhu, Z. Song, R. Chang, et al., A voxel-based morphometric study of age-and sex-related changes in white matter volume in the normal aging brain, Neuropsychiatr. Dis. Treat., 12 (2016), 453. |
| [15] |
N. Malykhin, S. Vahidy, S. Michielse, N. Coupland, R. Camicioli, P. Seres, et al., Structural organization of the prefrontal white matter pathways in the adult and aging brain measured by diffusion tensor imaging, Brain Struct. Funct., 216 (2011), 417-431. doi: 10.1007/s00429-011-0321-1
|
| [16] | L. Zhou, N. Tian, Z. J. Geng, B. K. Wu, L. Y. Dong, M. R. Wang, Diffusion tensor imaging study of brain precentral gyrus and postcentral gyrus during normal brain aging process, Brain Behav., 10 (2020), e01758. |
| [17] |
E. Bullmore, O. Sporns, Complex brain networks: graph theoretical analysis of structural and functional systems, Nat. Rev. Neurosci., 10 (2009), 186-198. doi: 10.1038/nrn2575
|
| [18] |
L. Deuker, E. T. Bullmore, M. Smith, S. Christensen, P. J. Nathan, B. Rockstroh, et al., Reproducibility of graph metrics of human brain functional networks, Neuroimage, 47 (2009), 1460-1468. doi: 10.1016/j.neuroimage.2009.05.035
|
| [19] | O. Sporns, G. Tononi, R. Kötter, The human connectome: a structural description of the human brain, PLoS Comput. Biol., 1 (2005), e42. |
| [20] |
H. Cheng, Y. Wang, J. Sheng, W. G. Kronenberger, V. P. Mathews, T. A. Hummer, et al., Characteristics and variability of structural networks derived from diffusion tensor imaging, Neuroimage, 61 (2012), 1153-1164. doi: 10.1016/j.neuroimage.2012.03.036
|
| [21] | O. Ajilore, M. Lamar, A. Kumar, Association of brain network efficiency with aging, depression, and cognition, Am. J. Geriatr. Psychiatry, 22 (2014), 102-110. |
| [22] | L. Lin, M. Tian, Q. Wang, S. Wu, Diffusion tensor tractography reveals disrupted structural connectivity during brain aging, in Journal of Physics: Conference Series, 910 (2017), 012042. |
| [23] |
T. Zhao, M. Cao, H. Niu, Z. Xi-Nian, A. Evans, Y. He, et al., Age-related changes in the topological organization of the white matter structural connectome across the human lifespan, Hum. Brain Mapp., 36 (2015), 3777-3792. doi: 10.1002/hbm.22877
|
| [24] |
N. A. Crossley, A. Mechelli, J. Scott, F. Carletti, P. T. Fox, P. McGuire, et al., The hubs of the human connectome are generally implicated in the anatomy of brain disorders, Brain, 137 (2014), 2382-2395. doi: 10.1093/brain/awu132
|
| [25] | C. Jin, L. Lin, L. W. Kuo, S. Wu, Z. Fu, Y. P. Chao, The relationships between the identified critical nodes within DTI-based brain structural network using hub measurements and vulnerability measurement, in 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), IEEE, (2015), 422-425. |
| [26] | W. Wen, R. Luo, X. Tang, L. Tang, H. X. Huang, X. Wen, et, al. Age-related progression of arterial stiffness and its elevated positive association with blood pressure in healthy people, Atherosclerosis, 238 (2015), 147-152. |
| [27] | Z. Cui, S. Zhong, P. Xu, Y. He, G. Gong, PANDA: a pipeline toolbox for analyzing brain diffusion images, Front. Hum. Neurosci., 7 (2013), 42. |
| [28] | G. Gong, Local diffusion homogeneity (LDH): an inter-voxel diffusion MRI metric for assessing inter-subject white matter variability, PLoS One, 8 (2013), e66366. |
| [29] |
W. Chau, A. R. McIntosh, The Talairach coordinate of a point in the MNI space: how to interpret it, Neuroimage, 25 (2005), 408-416. doi: 10.1016/j.neuroimage.2004.12.007
|
| [30] | J. Ashburner, SPM: a history, Neuroimage, 62 (2012), 791-800. |
| [31] | J. Klein, A. Grötsch, D. Betz, S. Barbieri, O. Friman, B. Stieltjes, et al., Qualitative and quantitative analysis of probabilistic and deterministic fiber tracking, in Medical Imaging 2010: Image Processing. International Society for Optics and Photonics, 7623 (2010), 76232A. |
| [32] |
S. Mori, Imaging cortical association using diffusion-tensor-based tracts in the human brain axonal tracking, Magn. Reson. Med., 47 (2002), 215-223. doi: 10.1002/mrm.10074
|
| [33] |
N. Tzourio-Mazoyer, B. Landeau, D. Papathanassiou, F. Crivello, O. Etard, N. Delcroix, et al., Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain, Neuroimage, 15 (2002), 273-289. doi: 10.1006/nimg.2001.0978
|
| [34] |
M. Rubinov, O. Sporns, Complex network measures of brain connectivity: uses and interpretations, Neuroimage, 52 (2010), 1059-1069. doi: 10.1016/j.neuroimage.2009.10.003
|
| [35] |
A. Zalesky, A. Fornito, I. H. Harding, L. Cocchi, M. Yücel, C. Pantelis, et al., Whole-brain anatomical networks: does the choice of nodes matter, Neuroimage, 50 (2010), 970-983. doi: 10.1016/j.neuroimage.2009.12.027
|
| [36] | A. Mantrach, L. Yen, J. Callut, K. Francoisse, M. Shimbo, M. Saerens, The sum-over-paths covariance kernel: A novel covariance measure between nodes of a directed graph, IEEE Trans. Pattern Anal. Mach. Intell., 32 (2009), 1112-1126. |
| [37] | F. U. Fischer, D. Wolf, A. Scheurich, A. Fellgiebel, Association of structural global brain network properties with intelligence in normal aging, PLoS One, 9 (2014), e86258. |
| [38] | Y. Sun, R. Lee, Y. Chen, S. Collinson, N. Thakor, A. Bezerianos, et al., Progressive gender differences of structural brain networks in healthy adults: a longitudinal, diffusion tensor imaging study, PLoS One, 10 (2015), e0118857. |
| [39] |
Y. F. Wang, P. Gu, J. Zhang, R. Qi, M. Veer, G. Zheng, et al. Deteriorated functional and structural brain networks and normally appearing functional-structural coupling in diabetic kidney disease: a graph theory-based magnetic resonance imaging study, Eur. Radiol., 29 (2019), 5577-5589. doi: 10.1007/s00330-019-06164-1
|
| [40] | J. Wang, X. Wang, M. Xia, X. Liao, A. Evans, Y. He, GRETNA: a graph theoretical network analysis toolbox for imaging connectomics, Front. Hum. Neurosci., 9 (2015), 386. |
| [41] | M. Xia, J. Wang, Y. He, BrainNet Viewer: a network visualization tool for human brain connectomics, PLoS One, 8 (2013), e68910. |
| [42] |
B. J. Cherry, M. Adamson, A. Duclos, J. B. Hellige, Aging and individual variation in interhemispheric collaboration and hemispheric asymmetry, Aging neuropsychol. C., 12 (2005), 316-339. doi: 10.1080/17444128.2005.10367004
|
| [43] | K. Wu, Y. Taki, K. Sato, H. Qi, R. Kawashima, H. Fukuda, A longitudinal study of structural brain network changes with normal aging, Front. Hum. Neurosci., 7 (2013), 113. |
| [44] |
Y. Liu, C. Yu, X. Zhang, J. Liu, Y. Duan, A. F. Alexander-Bloch, et al., Impaired long distance functional connectivity and weighted network architecture in Alzheimer's disease, Cereb. Cortex, 24 (2014), 1422-1435. doi: 10.1093/cercor/bhs410
|
| [45] |
G. Gong, Y. He, L. Concha, C. Lebel, D. W. Gross, A. C. Evans, et al., Mapping anatomical connectivity patterns of human cerebral cortex using in vivo diffusion tensor imaging tractography, Cereb. Cortex, 19 (2009), 524-536. doi: 10.1093/cercor/bhn102
|
| [46] |
B. M. Tijms, A. M. Wink, W. de Haan, W. M. van der Flier, C. J. Stam, P. Scheltens, et al., Alzheimer's disease: connecting findings from graph theoretical studies of brain networks, Neurobiol. Aging, 34 (2013), 2023-2036. doi: 10.1016/j.neurobiolaging.2013.02.020
|
| [47] | S. Xie, Z. Zhang, F. Chang, Y. Wang, Z. Zhang, Z. Zhou, et al., Subcortical white matter changes with normal aging detected by multi-shot high resolution diffusion tensor imaging, PLoS One, 11 (2016), e0157533. |
| [48] |
M. Di Paola, C. Caltagirone, G. Spalletta, What does the corpus callosum tell us about brain changes in the elderly, Expert Rev. Neurother., 11 (2011), 1557-1560. doi: 10.1586/ern.11.130
|

Figures(2) / Tables(5)

Song Xu, Xufeng Yao, Liting Han, Yuting Lv, Xixi Bu, Gan Huang, Yifeng Fan, Tonggang Yu, Gang Huang. Brain network analyses of diffusion tensor imaging for brain aging[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6066-6078. doi: 10.3934/mbe.2021303







 DownLoad:
DownLoad: