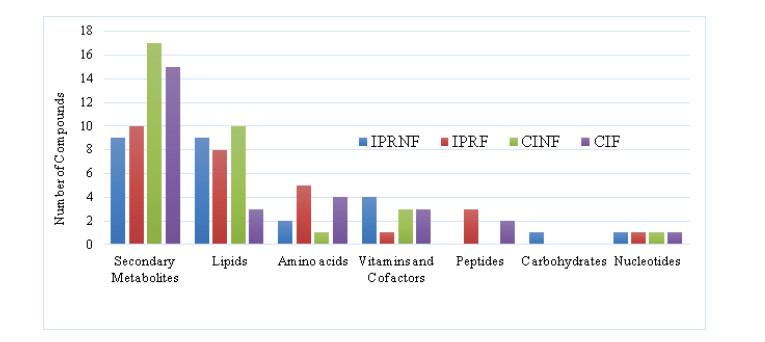
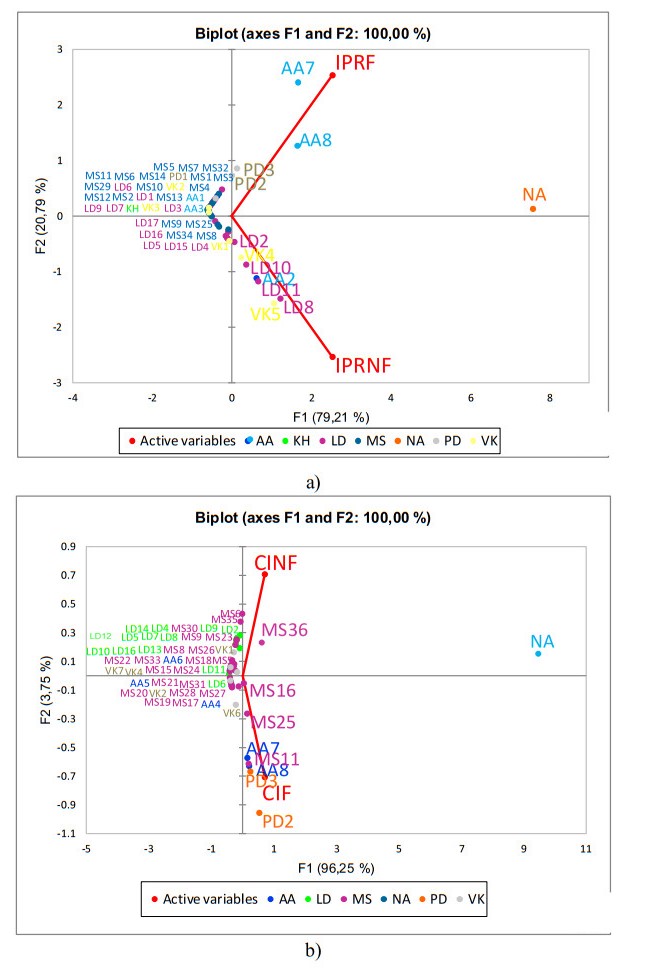
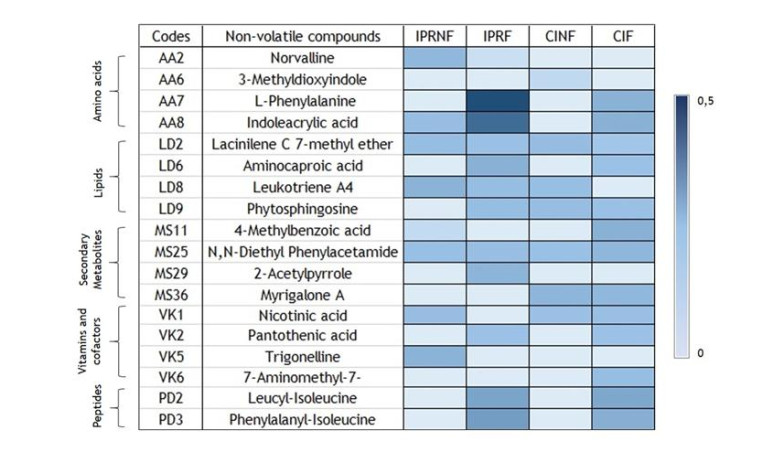
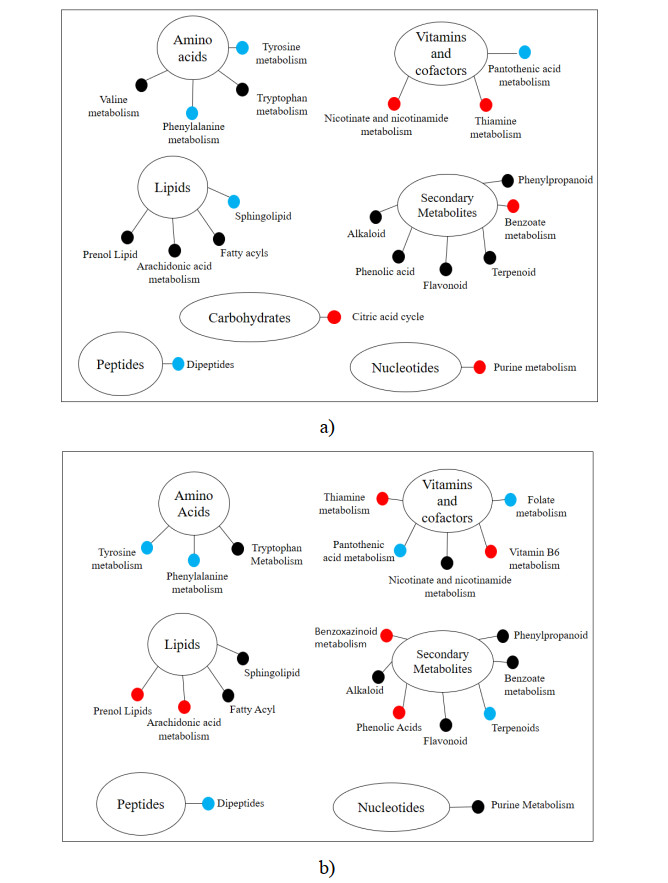
Rice bran, a by-product of the rice milling process, has high nutritional and bioactive components. The fermentation process has been adopted to enhance the nutritional quality of rice bran. The objective of this study was to identify and compare non-volatile compounds of fermented and non-fermented Inpari 30 and Cempo Ireng rice bran and its blood pressure-lowering activity. Each sample of rice bran was fermented by Rhizopous oligosporus for 72 hours at 30 ℃, using solid-state fermentation. In this study, non-fermented rice bran was used as a control. Non-targeted analysis of samples was carried out by Ultra performance liquid chromatography-tandem mass spectrometry with electrospray ionization in positive ion mode. For animal study, we used stroke-prone spontaneously hypertensive rats (SHRSP)-a model of hypertension-related disorders similar to human essential hypertension. Principal component analysis was done to categorize non-volatile compounds, based on its relative concentration. Seventy-two non-volatile compounds were identified in fermented and non-fermented rice bran, classified into secondary metabolites (36 compounds), lipids (16 compounds), amino acids (8 compounds), vitamins and cofactors (7 compounds), peptides (3 compounds), nucleotides (1 compound), and carbohydrates (1 compound). Adenosine was the most dominant of the non-volatile compounds in all rice bran samples. Further, blood pressure-lowering activity was shown in Inpari 30 fermented rice bran after six hours of administration in SHRSP. This study revealed a diversity of non-volatile compounds in fermented and non-fermented versions of Inpari 30 and Cempo Ireng rice bran that have anti-hypertensive effects. This study may provide opportunities to promote the production of fermented RB as a functional ingredient with enhanced bioactivity.
Citation: Ardiansyah, Fauziyyah Ariffa, Rizki Maryam Astuti, Wahyudi David, Dody Dwi Handoko, Slamet Budijanto, Hitoshi Shirakawa. Non-volatile compounds and blood pressure-lowering activity of Inpari 30 and Cempo Ireng fermented and non-fermented rice bran[J]. AIMS Agriculture and Food, 2021, 6(1): 337-359. doi: 10.3934/agrfood.2021021
Rice bran, a by-product of the rice milling process, has high nutritional and bioactive components. The fermentation process has been adopted to enhance the nutritional quality of rice bran. The objective of this study was to identify and compare non-volatile compounds of fermented and non-fermented Inpari 30 and Cempo Ireng rice bran and its blood pressure-lowering activity. Each sample of rice bran was fermented by Rhizopous oligosporus for 72 hours at 30 ℃, using solid-state fermentation. In this study, non-fermented rice bran was used as a control. Non-targeted analysis of samples was carried out by Ultra performance liquid chromatography-tandem mass spectrometry with electrospray ionization in positive ion mode. For animal study, we used stroke-prone spontaneously hypertensive rats (SHRSP)-a model of hypertension-related disorders similar to human essential hypertension. Principal component analysis was done to categorize non-volatile compounds, based on its relative concentration. Seventy-two non-volatile compounds were identified in fermented and non-fermented rice bran, classified into secondary metabolites (36 compounds), lipids (16 compounds), amino acids (8 compounds), vitamins and cofactors (7 compounds), peptides (3 compounds), nucleotides (1 compound), and carbohydrates (1 compound). Adenosine was the most dominant of the non-volatile compounds in all rice bran samples. Further, blood pressure-lowering activity was shown in Inpari 30 fermented rice bran after six hours of administration in SHRSP. This study revealed a diversity of non-volatile compounds in fermented and non-fermented versions of Inpari 30 and Cempo Ireng rice bran that have anti-hypertensive effects. This study may provide opportunities to promote the production of fermented RB as a functional ingredient with enhanced bioactivity.

| [1] | BPS (2019) Luas Panen dan Produksi Padi di Indonesia 2019. Badan Pusat Statistik. Jakarta: Badan Pusat Statistik 2019. Available from https://www.bps.go.id/publication/2019/07/04/daac1ba18cae1e90706ee58a/statistik-indonesia-2019.html. |
| [2] |
Oliveira Md, Feddern V, Kupski L, et al. (2010) Physico-chemical characterization of fermented rice bran biomass. Cyta-J Food 8: 229-236. doi: 10.1080/19476330903450274

|
| [3] |
Henderson AJ, Ollila CA, Kumar A, et al. (2012) Chemopreventive properties of dietary rice bran: current status and future prospects. Adv Nutr 3: 643-653. doi: 10.3945/an.112.002303
|
| [4] |
Ardiansyah, Ohsaki Y, Shirakawa H, et al. (2008) Novel effects of a single administration of ferulic acid on the regulation of blood pressure and the hepatic lipid metabolic profile in stroke-prone spontaneously hypertensive rats. J Agric Food Chem 56: 2825-2830. doi: 10.1021/jf072896y
|
| [5] |
Ardiansyah, Shirakawa H, Shimeno T, et al. (2009) Adenosine, an identified active component from the Driselase-treated fraction of rice bran, is effective at improving metabolic syndrome in stroke-prone spontaneously hypertensive rats. J Agric Food Chem 57: 2558-2564. doi: 10.1021/jf803347c
|
| [6] |
Lei L, Chen J, Liu Y, et al. (2018) Dietary wheat bran oil is equally effective as rice bran oil in reducing plasma cholesterol. J Agric Food Chem 66: 2765-2774. doi: 10.1021/acs.jafc.7b06093
|
| [7] |
Shao Y, Xu F, Sun X, et al. (2014) Identification and quantification of phenolic acids and anthocyanins as antioxidants in bran, embryo and endosperm of white, red and black rice kernels (Oryza sativa L.). J Cereal Sci 59: 211-218. doi: 10.1016/j.jcs.2014.01.004
|
| [8] | Kim D, Dong HG (2014) Fermented rice bran attenuates oxidative stress. In: Watson RS, Preedy VR, Zibadi S, Wheat and rice in disease prevention and health: benefits, risks and mechanisms of whole grains in health promotion, San Diego: Academic Press, 467-480. |
| [9] | Costa JA, Treichel H, Kumar V, et al. (2018) Advances in solid-state fermentation. In: Pandey A, Larroche C, Soccol CR, Current developments in biotechnology and bioengineering: current advances in solid state fermentation, Cambridge, MA, USA: Elsevier, 1-17. |
| [10] |
Razak DL, Rashid NY, Jamaluddin A, et al. (2014) Enhancement of phenolic acid content and antioxidant activity of rice bran fermented with Rhizopus oligosporus and Monascus purpureus. Biocatal Agric Biotechnol 4: 33-38. doi: 10.1016/j.bcab.2014.11.003
|
| [11] |
Ardiansyah, David W, Handoko DD, et al. (2019) Fermented rice bran extract improves blood pressure and glucose in stroke-prone spontaneously hypertensive rats. Nutr Food Sci 49: 844-853. doi: 10.1108/NFS-12-2018-0340
|
| [12] |
Zarei I, Brown DG, Nealon NJ, et al. (2017) Rice bran metabolome contains amino acids, vitamins & cofactors, and phytochemicals with medicinal and nutritional properties. Rice 10: 1-21. doi: 10.1186/s12284-017-0157-2
|
| [13] |
Zarei I, Luna E, McClung A, et al. (2018) Comparative rice bran metabolomics across diverse cultivars and functional rice gene-bran metabolite relationships. Metabolites 8: 1-22. doi: 10.3390/metabo8040063
|
| [14] |
Ryan EP, Heuberger AL, Weir TL, et al. (2011) Rice bran fermented with Saccharomyces boulardii generates novel metabolite profiles with bioactivity. J Agric Food Chem 59: 1862-1870. doi: 10.1021/jf1038103
|
| [15] | Ismail L, Osman R, Saim N (2013) Tandem solid phase extraction for the determination of pharmaceuticals in wastewater. Malaysian J Anal Sci 17: 262-271. |
| [16] |
Oikawa A, Otsuka T, Nakabayashi R, et al. (2015) Metabolic profiling of developing pear fruits reveals dynamic variation in primary and secondary metabolites, including plant. PLoS ONE 10: e0131408. doi: 10.1371/journal.pone.0131408
|
| [17] | Chen CD, Wan-Norafikah O, Lau K, et al. (2017) Laboratory study on the repellent bioefficacy of diethyl phenyl acetamide (DEPA) against Aedes aegypti. J Mosquito Res 7: 9-14. |
| [18] | Ito M, Shimura H, Watanabe N, et al. (1991) 2-Acetylpyrrole, a hepatoprotective compound from Streptomyces sp. A-5071. Agric Biol Chem 55: 2117-2121. |
| [19] |
Park C, Jeong NY, Kim G, et al. (2014) Momilactone B induces apoptosis and G1 arrest of the cell cycle in human monocytic leukemia U937 cells through downregulation of pRB phosphorylation and induction of the cyclin-dependent kinase inhibitor p21Waf1/Cip1. Oncol Rep 31: 1653-1660. doi: 10.3892/or.2014.3008
|
| [20] |
Quan NV, Tran HD, Xuan TD, et al. (2019) Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 24: 482. doi: 10.3390/molecules24030482
|
| [21] | Ammar HA, Ezzat SM, Houseny AM (2017) Improved production of kojic acid by mutagenesis of Aspergillus flavus HAk1 and Aspergillus oryzae HAk2 and their potential antioxidant activity. Biotech 7: 276. |
| [22] | Uzbekov VV, Talipov SA, Ibragimov BT, et al. (2013) Lacinilene C 7-methyl ether. Acta crystallographica. Section E, Structure reports online, 69. |
| [23] |
Kim S, Hong I, Hwang JS, et al. (2006) Phytosphingosine stimulates the differentiation of human keratinocytes and inhibits tpa-induced inflammatory epidermal hyperplasia in hairless mouse skin. Mol Med 12: 17-24. doi: 10.2119/2006-00001.Kim
|
| [24] |
Pavicic T, Wollenweber U, Farwick M, et al. (2007) Anti-microbial and -inflammatory activity and efficacy of phytosphingosine: an in vitro and in vivo study addressing acne vulgaris. Int J Cosmet Sci 181-190. doi: 10.1111/j.1467-2494.2007.00378.x
|
| [25] | Junne S, Klingner A, Itzeck D, et al. (2012) Consistency of Scale-Up from Bioprocess Development to Production. In: Subramanian G, Biopharmaceutical Production Technology. Wiley‐VCH Verlag GmbH & Co. KGaA, 511-543. |
| [26] | Bhagavan N (2015) Protein and amino acid metabolism. In: Bhagavan NV, Ha CE, Essentials of Medical Biochemistry, Toluca Lake, North Hollywood, CA, USA: Academic Press, 227-268. |
| [27] |
Zhou J, Zhou S, Zeng S (2011) Experimental diabetes treated with trigonelline: effect on b cell and pancreatic oxidative parameters. Fundam Clin Pharmacol 27: 279-287. doi: 10.1111/j.1472-8206.2011.01022.x
|
| [28] | Litwack G (2018) Chapter 20-Vitamins and nutrition. In: Litwack G, Human Biochemistry, Toluca Lake, North Hollywood, CA, USA: Academic Press, 645-680. |
| [29] |
Hamoud S, Kaplan M, Meilin E, et al. (2013) Niacin administration significantly reduces oxidative stress in patients with hypercholesterolemia and low levels of high-density lipoprotein cholesterol. Am J Med Sci 345: 195-199. doi: 10.1097/MAJ.0b013e3182548c28
|
| [30] |
Moura CS, Lollo PC, Morato PN, et al. (2017) Bioactivity of food peptides: biological response of rats to bovine milk whey peptides following acute exercise. Food Nutr Res 61: 1290740. doi: 10.1080/16546628.2017.1290740
|
| [31] |
Pokrovskiy MV, Korokin MV, Tsepeleva SA, et al. (2011) Arginase inhibitor in the pharmacological correction of endothelial dysfunction. Int J Hypertens 2011: 515047. doi: 10.4061/2011/515047
|
| [32] |
Wilza A, Pritchardb EM, Lia T, et al. (2008) Silk polymer based adenosine release: therapeutic potential for epileps. Biomaterials 29: 3609-3616. doi: 10.1016/j.biomaterials.2008.05.010
|
| [33] |
Williams-Karnesky, RL, Stenzel-Poore MP (2009) Adenosine and stroke: maximizing the therapeutic potential of adenosine as a prophylactic and acute neuroprotectant. Curr Neuropharmacol 7: 217-227. doi: 10.2174/157015909789152209
|
| [34] |
Sperlágh B, Vizi ES (2011) tTe role of extracellular adenosine in chemical neurotransmission in the hippocampus and basal ganglia: pharmacological and clinical aspects. Curr Top Med Chem 11: 1034-1046. doi: 10.2174/156802611795347564
|
| [35] |
Varzakas T (1998) Rhizopus oligosporus mycelial penetration and enzyme diffusion in soya bean tempe. Process Biochem 33: 741-747. doi: 10.1016/S0032-9592(98)00044-2
|
| [36] |
Ene IV, Brunke S, Brown AJ, et al. (2014) Metabolism in fungal pathogenesis. Cold Spring Harb Perspect Med 4: a019695. doi: 10.1101/cshperspect.a019695
|
| [37] |
Olsen I, Jantzen E (2001) Sphingolipids in bacteria and fungi. Anaerobe 7: 103-112. doi: 10.1006/anae.2001.0376
|
| [38] |
Oura T, Kajiwara, S (2010) Candida albicans sphingolipid C9-methyltransferase is involved in hyphal elongation. Microbiology 156: 1234-1243 doi: 10.1099/mic.0.033985-0
|
| [39] |
Epstein S, Castillon GA, Qin Y, et al. (2012) An essential function of sphingolipids in yeast cell division. Molecul Microbiol 84: 1018-1032. doi: 10.1111/j.1365-2958.2012.08087.x
|
| [40] | Schmidt-Dannert C (2015) Advances in Biochemical Engineering/Biotechnology. In: Jens S, Jorg B, Biotechnology of Isoprenoids, Switzerland: Springer, 19-61. |
| [41] |
Montagna DR, Duarte A, Todero MF, et al. (2020) Meta-tyrosine modulates the immune response induced by bacterial endotoxins. Immunobiology 225: 151856 doi: 10.1016/j.imbio.2019.10.005
|
| [42] |
Gueron G, Anselmino N, Chiarella P, et al. (2018) Game-changing restraint of ros-damaged phenylalanine, upon tumor metastasis. Cell Death Dis 9: 140. doi: 10.1038/s41419-017-0147-8
|
| [43] |
Hogan KL, Leiske D, Salisbury CM (2017) Characterization of N-Acetyl-Tryptophan degradation in protein therapeutic formulations. J Pharm Sci 106: 3499-3506. doi: 10.1016/j.xphs.2017.08.012
|
| [44] | Poewe W, Antonini A, Zijlmans JC, et al. (2010) Levodopa in the treatment of Parkinson's disease: an old drug still going strong. Clin Inter Aging 5: 229-238. |
| [45] |
Wlodarska M, Luo C, Kolde R, et al. (2017) Indoleacrylic acid produced by commensal peptostrepcoccus species suppresses inflamation. Cell Host Microbe 22: 25-37. doi: 10.1016/j.chom.2017.06.007
|
| [46] |
Kamzolova SV, Vinokurova NG, Yusupova AI, et al. (2012) Succinic acid production from n-alkanes. Eng Life Sci 12: 560-566. doi: 10.1002/elsc.201100241
|
| [47] |
Yihua LM, Jianwen, WM (2019) Efficacy of aminocaproic acid in the control of bleeding after total knee and hip arthroplast. Medicine 98: e14764. doi: 10.1097/MD.0000000000014764
|
| [48] | Strauss JF, FitzGerald GA (2014) Steroid hormones and other lipid molecules involved in human reproduction. In: Strauss JF, Barbieri RL, Yen & Jaffe's Reproductive Endocrinology (Seventh Edition), Cambridge, MA, USA: Elsevier, 66-92. |
| [49] |
Kang MS, Hirai S, Goto T, et al. (2008) Dehydroabietic acid, a phytochemical, acts as ligand for PPARs in macrophages and adipocytes to regulate inflammation. Biochem Biophy Res Commun 369: 333-338. doi: 10.1016/j.bbrc.2008.02.002
|
| [50] |
Kim J, Kang YG, Lee J, et al. (2015) The natural phytochemical dehydroabietic acid is an anti-aging reagent that mediates the direct activation of SIRT1. Mol Cell Endocrinol 412: 216-225. doi: 10.1016/j.mce.2015.05.006
|
| [51] |
Chang EYC, Chang YC, Shun CT, et al. (2016) Inhibition of prostaglandin reductase 2, a putative oncogene overexpressed in human pancreatic adenocarcinoma, induces oxidative stress-mediated cell death involving xCT and CTH gene expressions through 15-Keto-PGE2. PLoS One 11: e0147390. doi: 10.1371/journal.pone.0147390
|
| [52] |
Kanaoka Y, Austen KF (2019) Roles of cysteinyl leukotrienes and their receptors in immune cell-related functions. Advan Immunol 142: 65-84. doi: 10.1016/bs.ai.2019.04.002
|
| [53] |
Huang ZL, Urade Y, Hayaishi O (2011) The role of adenosine in the regulation of sleep. Curr Top Med Chem 11: 1047-1057. doi: 10.2174/156802611795347654
|
| [54] |
Seliger JM, Cicek SS, Witt LT, et al. (2018) Selective inhibition of human AKR1B10 by n-humulone, adhumulone and cohumulone isolated from Humulus lupulus extract. Molecules 23: 3041. doi: 10.3390/molecules23113041
|
| [55] | Silva GN, Pozzatti P, Rigatti F, et al. (2015) Antimicrobial evaluation of sesquiterpene α-curcumene and its synergism with imipenem. J Microbiol Biotechnol Food Sci 4: 434-436. |
| [56] |
Shin Y, Lee Y (2013) Cytotoxic activity from curcuma zedoaria through mitochondrial activation on ovarian cancer cells. Toxicol Res 29: 257-261. doi: 10.5487/TR.2013.29.4.257
|
| [57] | Rychen G, Aquilina G, Azimonti G, et al. (2018) Safety and efficacy of butylated hydroxyanisole (BHA) as a feed additive for all animal species. EFSA J 16: 1-18. |
| [58] | Baytas SN, Inceler N, Orhan DD, et al. (2011) Synthesis, characterization and antioxidant and antimicrobial properties of new ester and amide derivatives of indole-2-carboxylic acid. Fabad J Pharm Sci 36: 53-61. |
| [59] |
Saeedi M, Eslamifar M, Khezr, K (2019) Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed Pharmacother 110: 582-593. doi: 10.1016/j.biopha.2018.12.006
|
| [60] |
Kuete V, Metuno R, Ngameni B, et al (2007) Antimicrobial activity of the methanolic extracts and compounds from Treculia africana and Treculia acuminata (Moraceae). J Ethnopharmacol 112: 531-536. doi: 10.1016/j.jep.2007.04.010
|
| [61] |
Huang L, Su T, Li X (2013) Natural products as sources of new lead compounds for the treatment of Alzheimer's disease. Curr Top Med Chem 13: 1864-1878. doi: 10.2174/15680266113139990142
|
| [62] |
Gusdinar T, Herowati R, Kartasasmita R, et al. (2011) Anti-inflammatory and antioxidant activity of Quercetin-3, 3', 4'-Triacetate. J Pharmacol Toxicol 6: 182-188. doi: 10.3923/jpt.2011.182.188
|
| [63] |
Zandi K, Teoh BT, Sam SS, et al. (2011) Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol J 8: 560. doi: 10.1186/1743-422X-8-560
|
| [64] |
Edwards RL, Lyon T, Litwin SE, et al. (2007) Quercetin reduces blood pressure in hypertensive subjects. J Nutr 137: 2405-2411. doi: 10.1093/jn/137.11.2405
|
| [65] |
Vidyashankar S, Varma RS, Patki PS (2013) Quercetin ameliorate insulin resistance and up-regulates cellular antioxidants during oleic acid induced hepatic steatosis in HepG2 cells. Toxicol In Vitro 27: 945-953. doi: 10.1016/j.tiv.2013.01.014
|
| [66] | Choi JS, Jung MJ, Park HJ et al. (2002) Further Isolation of Peroxynitrite and 1, 1-Diphenyl-2-picrylhydrazyl Radical Scavenging Isorhamnetin 7-O-glucoside from the Leaves of Brassica juncea L. J Pharm Res 25: 625-627. |
| [67] |
Li W, Chen Z, Yan M, et al. (2016) The protective role of isorhamnetin on human brain microvascular endothelial cells from cytotoxicity induced by methylglyoxal and oxygen-glucose deprivation. J Neurochem 136: 651-659. doi: 10.1111/jnc.13436
|
| [68] |
Renugadevo J, Prabu SM (2010) Cadmium-induced hepatotoxicity in rat amd the protective. Exp Toxicol Pathol 62: 171-181. doi: 10.1016/j.etp.2009.03.010
|
| [69] |
Chen S, Ding Y, Tao W, et al. (2012) Naringenin inhibits TNF-α induced VSMC poliferation and migration via induction of HO-1. Food Chem Toxicol 50: 3025-3031. doi: 10.1016/j.fct.2012.06.006
|
| [70] | Rashmi R, Magesh SB, Ramkumar KM, et al. (2018) Antioxidant potential of naringenin helps to protect liver tissue from streptozotocin-induced damage. Rep Biochem Mol Biol 7: 76-84. |
| [71] |
Bawazeer NA, Choudary H, Zamzami MA, et al. (2016) Possible Regulation of LDL-Receptor by Naringenin in Hepg2 Hepatoma Cell Line. Afr J Tradit Complement Altern Med 14: 278-287. doi: 10.21010/ajtcam.v14i1.30
|
| [72] |
Frabasile S, Koishi AC, Kuczera D, et al. (2017) The citrus flavanone naringenin impairs dengue virus replication in human cells. Sci Rep 7: 41864. doi: 10.1038/srep41864
|
| [73] |
Lim W, Park S, Bazer FW, et al. (2017) Naringenin-induced apoptotic cell death in prostate cancer cells is mediated via the PI3K/AKT and MAPK signaling pathways. J Cell Biochem 118: 1118-1131. doi: 10.1002/jcb.25729
|
| [74] |
Haustein K (1999) Pharmacokinetic and pharmacodynamic properties of oral anticoagulants, especially phenprocoumon. Semin Thromb Hemostasis 25: 5-11. doi: 10.1055/s-2007-996417
|
| [75] |
Saroglou V, Karioti A, Rancic A, et al. (2009) Sesquiterpene lactones from anthemis melanolepis and their antibacterial and cytotoxic activities: prediction of their pharmacokinetic profile. J Nat Prod 73: 242-246. doi: 10.1021/np9004129
|
| [76] |
Tung YT, Wu JH, Kuo YH, et al. (2007) Antioxidant activities of natural phenolic compounds from Acacia confusa bark. Biores Tech 98: 1120-1123. doi: 10.1016/j.biortech.2006.04.017
|
| [77] | Nascimento GG, Locatelli J, Freitas P, et al. (2000) Antibacterial activity of plant extracts and phytochemicals on antibioticresistant bacteria. Braz J Microbiol 31: 247-256. |
| [78] | Kono Y, Uzawa J, Kobayashi K, et al. (1991) Structures of oryzalides A and B, and oryzalic acid A, a group of novel antimicrobial diterpenes, isolated from healthy leaves of a bacterial leaf blight-resistant cultivar of rice plants. Agric Biol Chem 55: 803-811. |
| [79] |
Killeen DP, Larsen L, Dayan FE, et al. (2016) Nortriketones: antimicrobial trimethylated acylphloroglucinols from Manuka (Leptospermum scoparium). J Nat Prod 79: 564-569. doi: 10.1021/acs.jnatprod.5b00968
|
| [80] | Miller JW, Rucker RB (2012) Panthothenic acid. In: Erdman W, Macdonald IA, Zeisel SH, Present knowledge in nutrition, 10th Edt Wiley-Blackwell, 375-389. |
| [81] |
Arlt A, Sebens S, Krebs S, et al. (2012) Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 32: 4825-4535. doi: 10.1038/onc.2012.493
|
| [82] |
Antonisamy P, Arasu MV, Dhanasekaran M, et al. (2015) Protective effects of trigonelline against indomethacin-induced gastric ulcer in rats and potential underlying mechanisms. Food Func 7: 398-408. doi: 10.1039/C5FO00403A
|
| [83] |
Fahanik-Babaei J, Baluchnejadmojarad T, Nikbakht F, et al. (2018) Trigonelline protects hippocampus against intracerebral Aβ(1-40) as a model of Alzheimer's disease in the rat: insights into underlying mechanisms. Metab Brain Dis 34: 191-201. doi: 10.1007/s11011-018-0338-8
|
| [84] |
McNulty H, Pentieva K, Hoey L, et al. (2008) Homocysteine, B-vitamins B and CVD. Proc Nutr Soc 67: 232-237. doi: 10.1017/S0029665108007076
|
| [85] | Boylan LM, Hart S, Porter KB, et al (2002) Vitamin B-6 content of breast milk and neonatal behavioral functioning. J Acad Nutr Diet 102: 1433-1438. |
| [86] |
Moura CS, Lollo PC, Morato PN, et al (2016) Functional effects of milk bioactive peptides on skeletal muscle of rats. Food Res Int 84: 18-25. doi: 10.1016/j.foodres.2016.03.001
|
Figures(4) / Tables(2)

Ardiansyah, Fauziyyah Ariffa, Rizki Maryam Astuti, Wahyudi David, Dody Dwi Handoko, Slamet Budijanto, Hitoshi Shirakawa. Non-volatile compounds and blood pressure-lowering activity of Inpari 30 and Cempo Ireng fermented and non-fermented rice bran[J]. AIMS Agriculture and Food, 2021, 6(1): 337-359. doi: 10.3934/agrfood.2021021







 DownLoad:
DownLoad: