Citation: Jinna Lu, Xiaoguang Zhang. Bifurcation analysis of a pair-wise epidemic model on adaptive networks[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2973-2989. doi: 10.3934/mbe.2019147

| [1] | F. Brauer and C. Castillo-Chavez, Mathematical models in population biology and epidemiology, New York: Springer, 2001. |
| [2] | R. Pastor-Satorras and A. Vespignani, Epidemic dynamics and endemic states in complex networks, Physic. Rev. E, 63 (2001), 649–667. |
| [3] | R. Pastor-Satorras and A. Vespignani, Epidemic dynamics in finite size scale-free networks, Physic. Rev. E, 65 (2002), 035108. |
| [4] | M. Boguñá and R. Pastor-Satorras, Epidemic spreading in correlated complex networks, Physic. Rev. E, 66 (2002), 047104. |
| [5] | P. V. Mieghem, J. Omic and R. Kooij, Virus spread in networks, IEEE ACM T. Network, 17 (2009), 1–14. |
| [6] | L. X. Yang, M. Draief and X. F. Yang, Heterogeneous virus propagation in networks: a theoretical study, Math. Method Appl. Sci , 40 (2017), 1396–1413. |
| [7] | L. X. Yang, X. F. Yang and Y. Wu, The impact of patch forwarding on the prevalence of computer virus: A theoretical assessment approach, Appl. Math. Model, 43 (2017), 110–125. |
| [8] | L. X. Yang, X. F. Yang and Y. Y. Tang, A Bi-Virus Competing Spreading Model with Generic Infection Rates, IEEE Transact. Netw. Sci. Eng., 5 (2018), 2–13. |
| [9] | X. Zhang, G. Q. Sun, Y. X. Zhu, et al., Epidemic dynamics on semi-directed complex networks, Math. Biosci., 246 (2013), 242–251. |
| [10] | J. Y. Yang, Y. M. Chen and F. Xu, Efect of infection age on an SIS epidemic model on complex networks, J. Math. Biol., 73 (2016), 1227–1249. |
| [11] | X. L. Peng, Z. Q. Zhang, J. Y. Yang, et al., An SIS epidemic model with vaccination in a dynamical contact network of mobile individuals with heterogeneous spatial constraints, Commun. Nonlinear Sci., 73 (2019), 52–73. |
| [12] | G. Q. Sun, C. H. Wang, L. L. Chang, et al., Efects of feedback regulation on vegetation patterns in semi-arid environments, Appl. Math. Model., 61 (2018), 200–215. |
| [13] | G. Q. Sun, M. Jusup, Z. Jin, et al., Pattern transitions in spatial epidemics: Mechanisms and emergent properties, Phys. Life Rev., 19 (2016), 43–73. |
| [14] | M. J. Keeling, D. A. Rand and A. J. Morris, Correlation models for childhood epidemics, P. Roy. Soc. B-Biol. Sci., 264 (1385), 1149–1156. |
| [15] | M. J. Keeling, The efects of local spatial structure on epidemiological invasions, Proc. R. Soc. Lond. B., 266 (1999), 859–867. |
| [16] | K. T. D. Eames and M. J. Keeling, Monogamous networks and the spread of sexually transmitted diseases, Math. Biosci., 189 (2004), 115–130. |
| [17] | Y.Wang, J. D. Cao, A. Alsaedi, et al., Edge-based SEIR dynamics with or without infectious force in latent period on random networks, Commun. Nonlinear Sci., 45 (2017), 35–54. |
| [18] | Y. Wang, J. D. Cao, X. D. Li, et al., Edge-based epidemic dynamics with multiple routes of transmission on random networks, Nonlinear Dynam., 91 (2018), 403–420. |
| [19] | Y.Wang, J. D. Cao and M. Q. Li, et al., Global behavior of a two-stage contact process on complex networks, J. Franklin I., In Press. |
| [20] | J. Y. Yang and F. Xu, The coumputational approach for the basic reproduction number of epidemic models on complex networks, IEEE Access, 2019, 10.1109/ACESS2019.2898639. |
| [21] | L. Li, C. H. Wang, S. F. Wang, et al., Hemorrhagic fever with renal syndrome in China: Mechanisms on two distinct annual peaks and control measures, Int. J. Biomath., 11 (2018), 1850030. |
| [22] | L. Li, J. Zhang, C. Liu, et al., Analysis of transmission dynamics for Zika virus on networks, Appl. Math. Comput., 347 (2019), 566–577. |
| [23] | D. J.Watts and S. H. Strogatz, Collective dynamics of 'small-world' networks, Nature, 393 (1998), 440–442. |
| [24] | A. L. Barabási and R. Albert, Emergence of scaling in random networks, Science, 286 (1999), 509–512. |
| [25] | R. M. May and A. L. Lloyd, Infection dynamics on scale-free networks, Physic. Rev. E, 64 (2001), 066112. |
| [26] | V. M. Eguiluz and K. Klemm, Epidemic threshold in structured scale-free networks, Physic. Rev. Lett., 89 (2002), 108701. |
| [27] | M. Boguna, R. Pastor-Satorras, A. Vespignani, Absence of epidemic threshold in scale-free networks with degree correlations, Physic. Rev. Lett., 90 (2003), 028701. |
| [28] | H. Zhang, P. Shu, Z.Wang, et al., Preferential imitation can invalidate targeted subsidy policies on seasonal-influenza diseases, Appl. Math. Comput., 294 (2017), 332–342. |
| [29] | T. Gross, C. J. D. D'Lima and B. Blasius, Epidemic dynamics on an adaptive network, Physic. Rev. Lett., 96 (2006), 208701. |
| [30] | T. Gross and B. Blasius, Adaptive coevolutionary networks: a review, J. R. Soc. Inter., 5 (2008), 259–271. |
| [31] | X. Zhang, C. Shan, Z. Jin, et al., Complex dynamics of epidemic models on adaptive networks, J. Difer. Equa., 266 (2019), 803–832. |
| [32] | L. B. Shaw and I. B. Schwartz, Fluctuating epidemics on adaptive networks, Physic. Rev. E, 77 (2008), 066101. |
| [33] | D. H. Zanette and S. Risau-Gusman, Infection Spreading in a Population with Evolving Contacts, J. Biol. Phys., 34 (2008), 135–148. |
| [34] | Y. Schwarzkopf, A. Ráos and D. Mukamel, Epidemic spreading in evolving networks, Physic. Rev. E, 82 (2010), 036112. |
| [35] | T. Rogers, W. Cliford-Brown, C. Mills, et al., Stochastic oscillations of adaptive networks: application to epidemic modelling, J. Stat. Mech. Theory. E, 8 (2012), 1–15. |
| [36] | D. Juher, J. Ripoll and J. Saldaña, Outbreak analysis of an SIS epidemic model with rewiring, J. Math. Biol., 67 (2013), 411–432. |
| [37] | J. Zhou, G. Xiao, S. A. Cheong, et al., Epidemic reemergence in adaptive complex networks, Physic. Rev. E, 85 (2012), 036107. |
| [38] | A. Szabó-Solticzky, L. Berthouze, I. Z. Kiss, et al., Oscillating epidemics in a dynamic network model: stochastic and mean-field analysis, J. Math. Biol., 72 (2016), 1153–1176. |
| [39] | S.J. Fan, A new extracting formula and a new distinguishing means on the one variable cubic equation, Nat. Sci. J. Hainan Teacheres Coll., 2 (1989), 91–98 (in Chinese). |
| [40] | S. N. Chow and J. K. Hale, Methods of bifurcation theory, Springer Science & Business Media, 2012. |
| [41] | L. Perko, Diferential equations and dynamical systems, Springer Science & Business Media, 2013. |
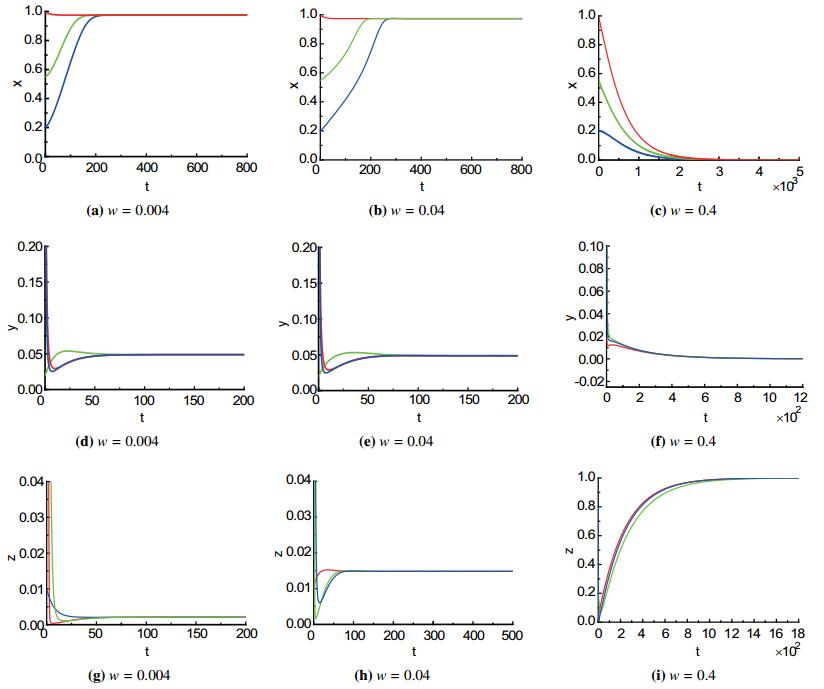
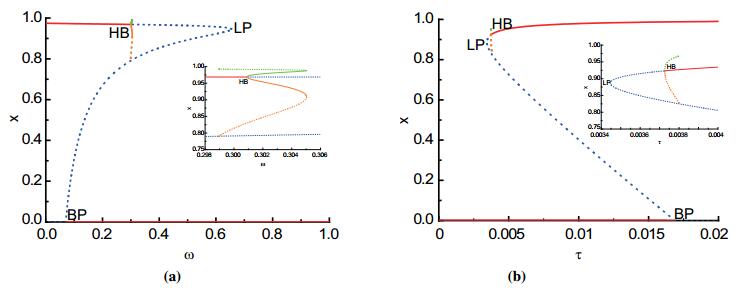
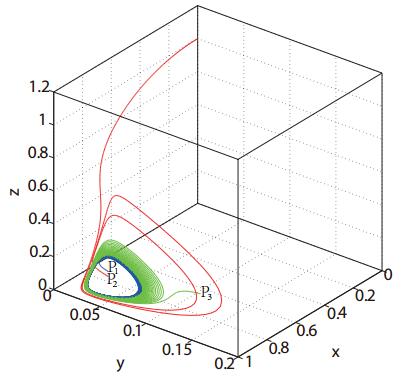
Figures(3)

Jinna Lu, Xiaoguang Zhang. Bifurcation analysis of a pair-wise epidemic model on adaptive networks[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2973-2989. doi: 10.3934/mbe.2019147







 DownLoad:
DownLoad: