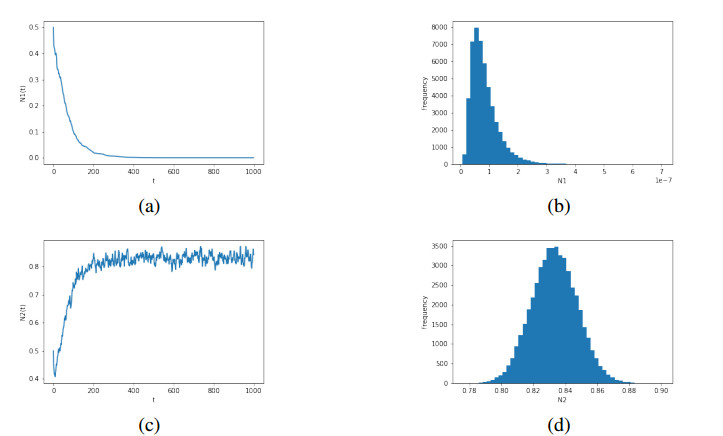
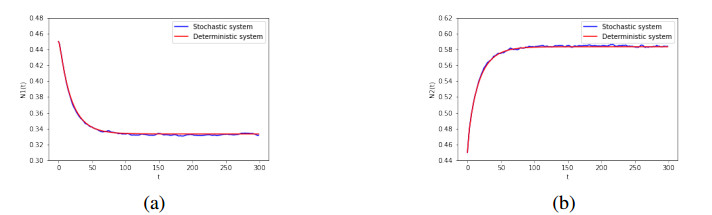
The competitive relationship is one of the important studies in population ecology. In this paper, we investigate the dynamical behaviors of a two-species Lotka-Volterra competition system in which intrinsic rates of increase are governed by the Ornstein-Uhlenbeck process. First, we prove the existence and uniqueness of the global solution of the model. Second, the extinction of populations is discussed. Moreover, a sufficient condition for the existence of the stationary distribution in the system is obtained, and, further, the formulas for the mean and the covariance of the probability density function of the corresponding linearized system near the equilibrium point are obtained. Finally, numerical simulations are applied to verify the theoretical results.
Citation: Huili Wei, Wenhe Li. Dynamical behaviors of a Lotka-Volterra competition system with the Ornstein-Uhlenbeck process[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 7882-7904. doi: 10.3934/mbe.2023341
The competitive relationship is one of the important studies in population ecology. In this paper, we investigate the dynamical behaviors of a two-species Lotka-Volterra competition system in which intrinsic rates of increase are governed by the Ornstein-Uhlenbeck process. First, we prove the existence and uniqueness of the global solution of the model. Second, the extinction of populations is discussed. Moreover, a sufficient condition for the existence of the stationary distribution in the system is obtained, and, further, the formulas for the mean and the covariance of the probability density function of the corresponding linearized system near the equilibrium point are obtained. Finally, numerical simulations are applied to verify the theoretical results.

| [1] |
V. Volterra, Variations and fluctuations of the number of individuals in animal species living together, ICES J. Mar. Sci., 3 (1928), 3–51. https://doi.org/10.1093/icesjms/3.1.3 doi: 10.1093/icesjms/3.1.3

|
| [2] |
N. Abdellatif, R. Fekih-Salem, T. Sari, Competition for a single resource and coexistence of several species in the chemostat, Math. Biosci. Eng., 13 (2016), 631. https://doi.org/10.3934/mbe.2016012 doi: 10.3934/mbe.2016012
|
| [3] |
B. S. Han, Z. C. Wang, Turing patterns of a Lotka-Volterra competitive system with nonlocal delay, Int. J. Bifurcation Chaos, 28 (2018), 1830021. https://doi.org/10.1142/S0218127418300215 doi: 10.1142/S0218127418300215
|
| [4] |
M. K. A. Gavina, T. Tahara, K. Tainaka, H. Ito, S. Morita, G. Ichinose, et al., Multi-species coexistence in Lotka-Volterra competitive systems with crowding effects, Sci. Rep., 8 (2018), 1–8. https://doi.org/10.1038/s41598-017-19044-9 doi: 10.1038/s41598-017-19044-9
|
| [5] |
D. Q. Jiang, C. Y. Ji, X. Y. Li, D. O'Regand, Analysis of autonomous Lotka-Volterra competition systems with random perturbation, J. Math. Anal. Appl., 390 (2012), 582–595. https://doi.org/10.1016/j.jmaa.2011.12.049 doi: 10.1016/j.jmaa.2011.12.049
|
| [6] |
K. Golpalsamy, Globally asymptotic stability in a periodic Lotka-Volterra system, J. Aust. Math. Soc. Ser. B, 27 (1982), 66–72. https://doi.org/10.1017/S0334270000004768 doi: 10.1017/S0334270000004768
|
| [7] |
Y. S. Wang, H. Wu, D. L. DeAngelis, Global dynamics of a mutualism-competition model with one resource and multiple consumers, J. Math. Biol., 78 (2019), 683–710. https://doi.org/10.1007/s00285-018-1288-9 doi: 10.1007/s00285-018-1288-9
|
| [8] |
Z. A. Wang, J. Xu, On the Lotka-Volterra competition system with dynamical resources and density-dependent diffusion, J. Math. Biol., 82 (2021), 1–37. https://doi.org/10.1007/s00285-021-01562-w doi: 10.1007/s00285-021-01562-w
|
| [9] |
R. M. May, Biological populations with nonoverlapping generations: stable points, stable cycles, and chaos, Science, 186 (1974), 645–647. https://doi.org/10.1126/science.186.4164.645 doi: 10.1126/science.186.4164.645
|
| [10] |
C. J. Mode, M. E. Jacobson, A study of the impact of environmental stochasticity on extinction probabilities by Monte Carlo integration, Math. Biosci., 83 (1987), 105–125. https://doi.org/10.1016/0025-5564(87)90006-X doi: 10.1016/0025-5564(87)90006-X
|
| [11] |
P. J. DuBowy, Waterfowl communities and seasonal environments: temporal variability in interspecific competition, Ecology, 69 (1988), 1439–1453. https://doi.org/10.2307/1941641 doi: 10.2307/1941641
|
| [12] |
X. Y. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stoch. Proc. Appl., 97 (2002), 95–110. https://doi.org/10.1016/S0304-4149(01)00126-0 doi: 10.1016/S0304-4149(01)00126-0
|
| [13] |
A. Babaei, H. Jafari, S. Banihashemi, M. Ahmadi, Mathematical analysis of a stochastic model for spread of Coronavirus, Chaos, Solitons Fractals, 145 (2021), 110788. https://doi.org/10.1016/j.chaos.2021.110788 doi: 10.1016/j.chaos.2021.110788
|
| [14] |
D. Li, J. A. Cui, M. Liu, S. Q. Liu, The evolutionary dynamics of stochastic epidemic model with nonlinear incidence rate, Bull. Math. Biol., 77 (2015), 1705–1743. https://doi.org/10.1007/s11538-015-0101-9 doi: 10.1007/s11538-015-0101-9
|
| [15] | D. J. Wilkinson, Stochastic Modelling for Systems Biology, 3$^{nd}$ edition, Chapman and Hall/CRC, 2018. https://doi.org/10.1201/9781351000918 |
| [16] |
F. F. Zhu, X. Z. Meng, T. H. Zhang, Optimal harvesting of a competitive n-species stochastic model with delayed diffusions, Math. Biosci. Eng., 16 (2019), 1554–1574. https://doi.org/10.3934/mbe.2019074 doi: 10.3934/mbe.2019074
|
| [17] |
R. Zhang, J. L. Wang, S. Q. Liu, Traveling wave solutions for a class of discrete diffusive SIR epidemic model, J. Nonlinear Sci., 31 (2021), 1–33. https://doi.org/10.1007/s00332-020-09656-3 doi: 10.1007/s00332-020-09656-3
|
| [18] |
S. He, S. Y. Tang, L. B. Rong, A discrete stochastic model of the COVID-19 outbreak: Forecast and control, Math. Biosci. Eng., 17 (2020), 2792–2804. https://doi.org/10.3934/mbe.2020153 doi: 10.3934/mbe.2020153
|
| [19] |
E. Allen, Environmental variability and mean-reverting processes, Discrete Cont. Dyn.-B, 21 (2016), 2073. https://doi.org/10.3934/dcdsb.2016037 doi: 10.3934/dcdsb.2016037
|
| [20] |
X. F. Zhang, R. Yuan, A stochastic chemostat model with mean-reverting Ornstein-Uhlenbeck process and Monod-Haldane response function, Appl. Math. Comput., 394 (2021), 125833. https://doi.org/10.1016/j.amc.2020.125833 doi: 10.1016/j.amc.2020.125833
|
| [21] |
Y. Q. Song, X. H. Zhang, Stationary distribution and extinction of a stochastic SVEIS epidemic model incorporating Ornstein-Uhlenbeck process, Appl. Math. Lett., 133 (2022), 108284. https://doi.org/10.1016/j.aml.2022.108284 doi: 10.1016/j.aml.2022.108284
|
| [22] |
Q. Yang, X. H. Zhang, D. Q. Jiang, Dynamical behaviors of a stochastic food chain system with Ornstein-Uhlenbeck process, J. Nonlinear Sci., 32 (2022), 1–40. https://doi.org/10.1007/s00332-022-09796-8 doi: 10.1007/s00332-022-09796-8
|
| [23] |
G. Ascione, Y. Mishura, E. Pirozzi, Fractional Ornstein-Uhlenbeck process with stochastic forcing, and its applications, Methodol. Comput. Appl. Probab., 23 (2021), 53–84. https://doi.org/10.1007/s11009-019-09748-y doi: 10.1007/s11009-019-09748-y
|
| [24] |
W. M. Wang, Y. L. Cai, Z. Q. Ding, Z. J. Gui, A stochastic differential equation SIS epidemic model incorporating Ornstein-Uhlenbeck process, Phys. A, 509 (2018), 921–936. https://doi.org/10.1016/j.physa.2018.06.099 doi: 10.1016/j.physa.2018.06.099
|
| [25] |
W. R. Li, Q. M. Zhang, M. B. Anke, M. Ye, Y. Li, Taylor approximation of the solution of age-dependent stochastic delay population equations with Ornstein-Uhlenbeck process and Poisson jumps, Math. Biosci. Eng., 17 (2020), 2650–2675. https://doi.org/10.3934/mbe.2020145 doi: 10.3934/mbe.2020145
|
| [26] |
X. F. Zhang, A stochastic non-autonomous chemostat model with mean-reverting Ornstein-Uhlenbeck process on the washout rate, J. Dyn. Differ. Equations, 2022. https://doi.org/10.1007/s10884-022-10181-y doi: 10.1007/s10884-022-10181-y
|
| [27] |
Y. A. Zhou, D. Q. Jiang, Dynamical behavior of a stochastic SIQR epidemic model with Ornstein-Uhlenbeck process and standard incidence rate after dimensionality reduction, Commun. Nonlinear Sci., 116 (2022), 106878. https://doi.org/10.1016/j.cnsns.2022.106878 doi: 10.1016/j.cnsns.2022.106878
|
| [28] |
Y. L. Cai, J. J. Jiao, Z. J. Gui, Y. T. Liu, W. M. Wang, Environmental variability in a stochastic epidemic model, Appl. Math. Comput., 329 (2018), 210–226. https://doi.org/10.1016/j.amc.2018.02.009 doi: 10.1016/j.amc.2018.02.009
|
| [29] |
A. Gray, D. Greenhalgh, L. Hu, X. Mao, J. Pan, A stochastic differential equation SIS epidemic model, SIAM J. Appl. Math., 71 (2011), 876–902. https://doi.org/10.1137/10081856X doi: 10.1137/10081856X
|
| [30] |
A. O. Akdemir, S. I. Butt, M. Nadeem, M. A. Ragusa, New general variants of Chebyshev type inequalities via generalized fractional integral operators, Mathematics, 9 (2021), 122. https://doi.org/10.3390/math9020122 doi: 10.3390/math9020122
|
| [31] |
H. P. Liu, Z. E. Ma, The threshold of survival for system of two species in a polluted environment, J. Math. Biol., 30 (1991), 49–61. https://doi.org/10.1007/BF00168006 doi: 10.1007/BF00168006
|
| [32] | R. Khasminskii, Stochastic Stability of Differential Equations, 2$^{nd}$ edition, Springer Berlin, Heidelberg, 2011. https://doi.org/10.1007/978-3-642-23280-0 |
| [33] |
R. Mahardika, Y. D. Sumanto, Routh-hurwitz criterion and bifurcation method for stability analysis of tuberculosis transmission model, J. Phys.: Conf. Ser., 1217 (2019), 012056. https://doi.org/10.1088/1742-6596/1217/1/012056 doi: 10.1088/1742-6596/1217/1/012056
|
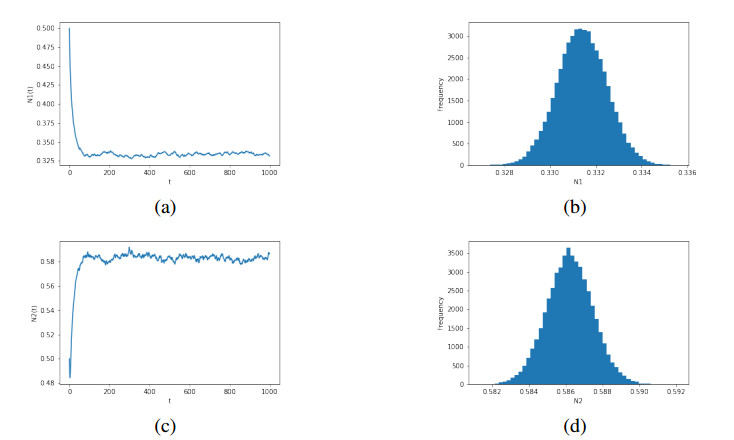
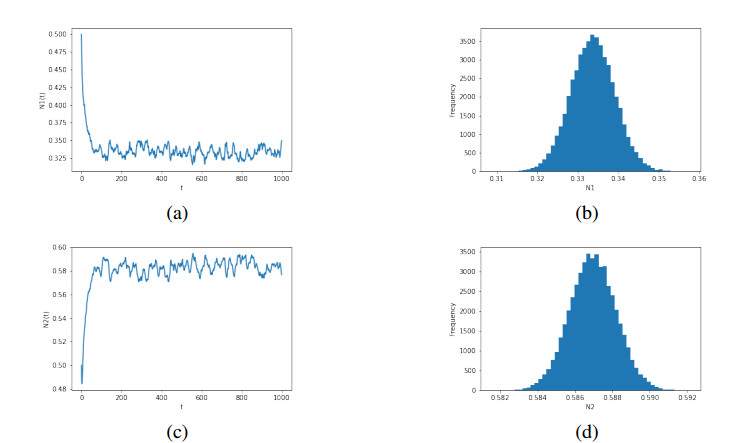
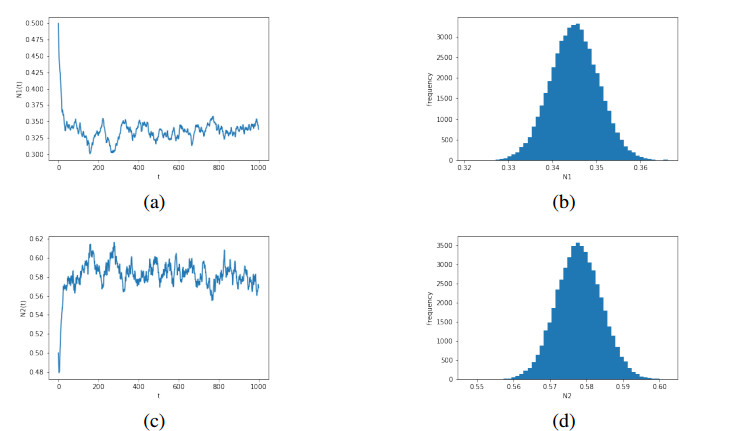
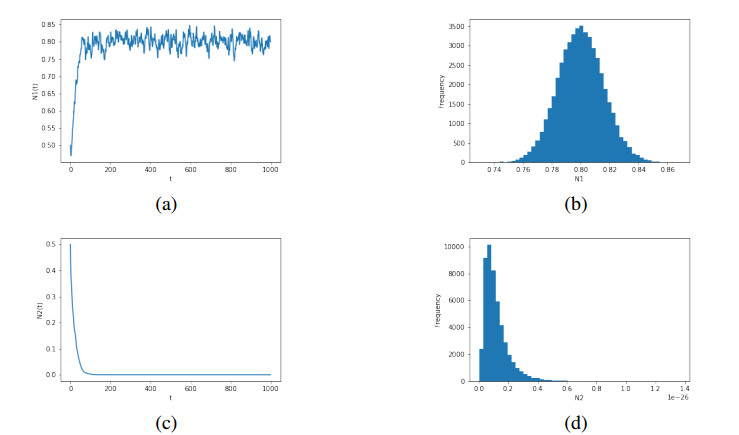
Figures(6)

Huili Wei, Wenhe Li. Dynamical behaviors of a Lotka-Volterra competition system with the Ornstein-Uhlenbeck process[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 7882-7904. doi: 10.3934/mbe.2023341







 DownLoad:
DownLoad: