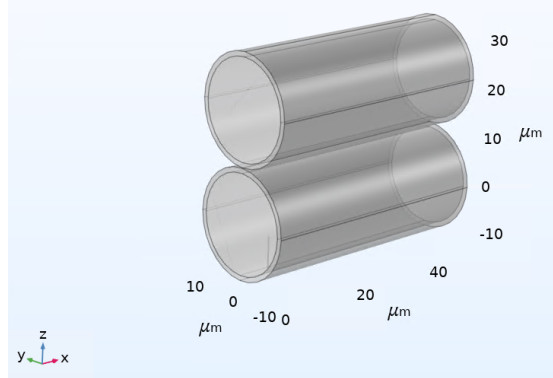
We have developed a numerical model of two osculating cylindrical elastic renal tubules to investigate the impact of neighboring tubules on the stress applied to a primary cilium. We hypothesize that the stress at the base of the primary cilium will depend on the mechanical coupling of the tubules due to local constrained motion of the tubule wall. The objective of this work was to determine the in-plane stresses of a primary cilium attached to the inner wall of one renal tubule subject to the applied pulsatile flow, with a neighboring renal tube filled with stagnant fluid in close proximity to the primary tubule. We used the commercial software COMSOLⓇ to model the fluid-structure interaction of the applied flow and tubule wall, and we applied a boundary load to the face of the primary cilium during this simulation to produces a stress at its base. We confirm our hypothesis by observing that on average the in-plane stresses are greater at the base of the cilium when there is a neighboring renal tube versus if there is no neighboring tube at all. In combination with the hypothesized function of a cilium as a biological fluid flow sensor, these results indicate that flow signaling may also depend on how the tubule wall is constrained by neighboring tubules. Our results may be limited in their interpretation due to the simplified nature of our model geometry, and further improvements to the model may potentially lead to the design of future experiments.
Citation: Nerion Zekaj, Shawn D. Ryan, Andrew Resnick. Fluid-structure interaction modelling of neighboring tubes with primary cilium analysis[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 3677-3699. doi: 10.3934/mbe.2023172
We have developed a numerical model of two osculating cylindrical elastic renal tubules to investigate the impact of neighboring tubules on the stress applied to a primary cilium. We hypothesize that the stress at the base of the primary cilium will depend on the mechanical coupling of the tubules due to local constrained motion of the tubule wall. The objective of this work was to determine the in-plane stresses of a primary cilium attached to the inner wall of one renal tubule subject to the applied pulsatile flow, with a neighboring renal tube filled with stagnant fluid in close proximity to the primary tubule. We used the commercial software COMSOLⓇ to model the fluid-structure interaction of the applied flow and tubule wall, and we applied a boundary load to the face of the primary cilium during this simulation to produces a stress at its base. We confirm our hypothesis by observing that on average the in-plane stresses are greater at the base of the cilium when there is a neighboring renal tube versus if there is no neighboring tube at all. In combination with the hypothesized function of a cilium as a biological fluid flow sensor, these results indicate that flow signaling may also depend on how the tubule wall is constrained by neighboring tubules. Our results may be limited in their interpretation due to the simplified nature of our model geometry, and further improvements to the model may potentially lead to the design of future experiments.

| [1] | Robert A. Bloodgood, Chapter 1–from central to rudimentary to primary: The history of an underappreciated organelle whose time has come.the primary cilium, in Methods in Cell Biology, Academic Press. https://doi.org/10.1016/S0091-679X(08)94001-2 |
| [2] |
X. Jin, A. M. Mohieldin, B. S. Muntean, J. A. Green, J. V. Shah, K. Mykytyn, et al., Cilioplasm is a cellular compartment for calcium signaling in response to mechanical and chemical stimuli, Cell. Mol. Life Sci., 71 (2014), 2165–2178. https://doi.org/10.1007/s00018-013-1483-1 doi: 10.1007/s00018-013-1483-1

|
| [3] |
S. Nag, A. Resnick, Biophysics and biofluid dynamics of primary cilia: Evidence for and against the flow-sensing function, Am. J. Physiol. Renal Physiol., 313 (2017), F706–F720. https://doi.org/10.1152/ajprenal.00172.2017 doi: 10.1152/ajprenal.00172.2017
|
| [4] |
N. Praljak, S. D. Ryan, A. Resnick, Pulsatile flow through idealized renal tubules: Fluid-structure interaction and dynamic pathologies, Math. Biosci. Eng., 17 (2019), 1787–1807. https://doi.org/10.3934/mbe.2020094 doi: 10.3934/mbe.2020094
|
| [5] | C. Rouiller, 2-General anatomy and histology of the kidney, in The Kidney, Academic Press. https://doi.org/10.1016/B978-1-4832-2825-9.50008-5 |
| [6] |
A.K O'Connor, E. B. Malarkey, N. F. Berbari, M. J. Croyle, C. J. Haycraft, P. D. Bell, et al., An inducible CiliaGFP mouse model for in vivo visualization and analysis of cilia in live tissue, Cilia, 2013 (2013), 8. https://doi.org/10.1186/2046-2530-2-8 doi: 10.1186/2046-2530-2-8
|
| [7] |
L. M. Satlin, S. Sheng, C. B. Woda, T. R. Kleyman, Epithelial $Na^{+}$ channels are regulated by flow, Am. J. Physiol. Renal Physiol., 280 (2001), 1010–1018. https://doi.org/10.1152/ajprenal.2001.280.6.F1010 doi: 10.1152/ajprenal.2001.280.6.F1010
|
| [8] | H. A. Praetorius, K. R. Spring, The renal cell primary cilium functions as a flow sensor, Curr. Opin. Nephrol. Hypertens., 12 (2003), 517–520. |
| [9] |
I. Mnassri, A. El Baroudi, Vibrational frequency analysis of finite elastic tube filled with compressible viscous fluid, Acta Mech. Solida Sin., 30 (2017), 435–444. https://doi.org/10.1016/j.camss.2017.07.010 doi: 10.1016/j.camss.2017.07.010
|
| [10] |
O. San, A. E. Staples, Dynamics of pulsatile flows through elastic microtubes, Int. J. Appl. Mech., 4 (2012), 1250006. https://doi.org/10.1142/S175882511200135X doi: 10.1142/S175882511200135X
|
| [11] |
M. E. Downs, A. M. Nguyen, F. A. Herzog, D. A. Hoey, C. R. Jacobs, An experimental and computational analysis of primary cilia deflection under fluid flow, Comput. Methods Biomech. Biomed. Eng., 17 (2014), 2–10. https://doi.org/10.1080/10255842.2011.653784 doi: 10.1080/10255842.2011.653784
|
| [12] |
H. Khayyeri, S. Barreto, D. Lacroix, Primary cilia mechanics affects cell mechanosensation: A computational study, J. Theor. J., 379 (2015), 38–46. https://doi.org/10.1016/j.jtbi.2015.04.034 doi: 10.1016/j.jtbi.2015.04.034
|
| [13] |
S. Sun, R. L. Fisher, S. S. Bowser, B. T. Pentecost, H. Sui, Three-dimensional architecture of epithelial primary cilia, Proc. Natl. Acad. Sci., 116 (2019), 9370–9379. https://doi.org/10.1073/pnas.1821064116 doi: 10.1073/pnas.1821064116
|
| [14] |
B. K. Yoder, Role of primary cilia in the pathogenesis of polycystic kidney disease, J. Am. Soc. Nephrol., 18 (2007), 1381–1388. https://doi.org/10.1681/ASN.2006111215 doi: 10.1681/ASN.2006111215
|
| [15] |
S. Wang, Z. Dong, Primary cilia and kidney injury: current research status and future perspectives, Am. J. Physiol. Renal Physiol., 305 (2013), F1085–F1098. https://doi.org/10.1152/ajprenal.00399.2013 doi: 10.1152/ajprenal.00399.2013
|
| [16] |
Z. Peng, A. Resnick, Y. N. Young, Primary cilium: a paradigm for integrating mathematical modeling with experiments and numerical simulations in mechanobiology, Math. Biosci. Eng., 18 (2021), 1215–1237. https://doi.org/10.3934/mbe.2021066 doi: 10.3934/mbe.2021066
|
| [17] |
L. C. Espinha, D. A. Hoey, P. R. Fernandes, H. C. Rodrigues, C. R. Jacobs, Oscillatory fluid flow influences primary cilia and microtubule mechanics, Cytoskeleton, 71 (2014), 435–445. https://doi.org/10.1002/cm.21183 doi: 10.1002/cm.21183
|
| [18] |
T. Sakai, D. A. Craig, A. S. Wexler, D. J. Marsh. Fluid waves in renal tubules, Biophys. J., 50 (1986), 805–813. https://doi.org/10.1016/S0006-3495(86)83521-4 doi: 10.1016/S0006-3495(86)83521-4
|
| [19] |
N. H. Holstein-Rathlou, D. J. Marsh, Oscillations of tubular pressure, flow, and distal chloride concentration in rats, Am. J. Physiol. Renal Physiol., 256 (1989), F1007–F1014. https://doi.org/10.1152/ajprenal.1989.256.6.F1007 doi: 10.1152/ajprenal.1989.256.6.F1007
|
| [20] |
S. Cortell, F. J. Gennari, M. Davidman, W. H. Bossert, W. B. Schwartz, A definition of proximal and distal tubular compliance. practical and theoretical implications, J. Clin. Invest., 52 (1973), 2330–2339. https://doi.org/10.1172/JCI107422 doi: 10.1172/JCI107422
|
| [21] | COMSOL Multiphysics. Available from: www.comsol.com. |
| [22] |
J. Flaherty, Z. Feng, Z. Peng, Y. N. Young, A. Resnick, Primary cilia have a length-dependent persistence length, Biomech. Model. Mechanobiol., 19 (2020), 445–460. https://doi.org/10.1007/s10237-019-01220-7 doi: 10.1007/s10237-019-01220-7
|
| [23] |
S. Rydholm, G. Zwartz, J. M. Kowalewski, P. K. Zare, T. Frisk, H. Brismar, Mechanical properties of primary cilia regulate the response to fluid flow, Am. J. Physiol. Renal Physiol., 298 (2009), F1096–F1102. https://doi.org/10.1152/ajprenal.00657.2009 doi: 10.1152/ajprenal.00657.2009
|
| [24] |
P. S. Mathieu, J. C. Bodle, E. G. Loboa, Primary cilium mechanotransduction of tensile strain in 3d culture: Finite element analyses of strain amplification caused by tensile strain applied to a primary cilium embedded in a collagen matrix, J. Biomech., 47 (2014), 2211–2217. https://doi.org/10.1016/j.jbiomech.2014.04.004 doi: 10.1016/j.jbiomech.2014.04.004
|
| [25] |
J. Cui, T. Wu, Y. Liu, B. M. Fu, Y. Jin, Z. Zhu, A three-dimensional simulation of the dynamics of primary cilia in an oscillating flow, Appl. Math. Modell., 108 (2022), 825–839. https://doi.org/10.1016/j.apm.2022.04.024 doi: 10.1016/j.apm.2022.04.024
|
| [26] |
J. O'Connor, A. Revell, P. Mandal, P. Day, Application of a lattice boltzmann-immersed boundary method for fluid-filament dynamics and flow sensing, J. Biomech., 49 (2016), 2143–2151. https://doi.org/10.1016/j.jbiomech.2015.11.057 doi: 10.1016/j.jbiomech.2015.11.057
|
| [27] | COMSOL multiphysics, Fluid structure interaction. Available from: https://www.comsol.com/model/fluid-structure-interaction-361. |
| [28] | COMSOL multiphysics, Contact Analysis. Available from: https://doc.comsol.com/5.5/doc/com.comsol.help.sme/sme_ug_theory.06.65.html. |
| [29] |
H. P. Peters, C. M. Laarakkers, P. Pickkers, R. Masereeuw, O. C. Boerman, A. Eek, et al., Tubular reabsorption and local production of urine hepcidin-25, BMC Nephrol., 14 (2013), 70. https://doi.org/10.1186/1471-2369-14-70 doi: 10.1186/1471-2369-14-70
|
| [30] | J. Howard, R. L. Clark, Mechanics of motor proteins and the cytoskeleton, Appl. Mech. Rev., 55 (2002), B39. |
| [31] |
B. Vahidi, N. Fatouraee, A. Imanparast, A. N. Moghadam, A mathematical simulation of the ureter: Effects of the model parameters on ureteral pressure/flow relations, J. Biomech. Eng., 133 (2011), 031004. https://doi.org/10.1115/1.4003316 doi: 10.1115/1.4003316
|
| [32] |
M. Pradella, R. M. Dorizzi, F. Rigolin, B. E. Statland, Relative density of urine: methods and clinical significance, Crit. Rev. Clin. Lab. Sci., 26 (1988), 195–242. https://doi.org/10.3109/10408368809105890 doi: 10.3109/10408368809105890
|
| [33] | C. Battle, Mechanics & dynamics of the primary cilium, Ph. D thesis, Georg-August-University G''ottingen, 2013. http://dx.doi.org/10.53846/goediss-4037 |
| [34] | COMSOL multiphysics, Meshing for Contact Analysis. Available from: https://doc.comsol.com/5.5/doc/com.comsol.help.sme/sme_ug_modeling.05.102.html. |

















Figures(18) / Tables(1)

Nerion Zekaj, Shawn D. Ryan, Andrew Resnick. Fluid-structure interaction modelling of neighboring tubes with primary cilium analysis[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 3677-3699. doi: 10.3934/mbe.2023172







 DownLoad:
DownLoad: