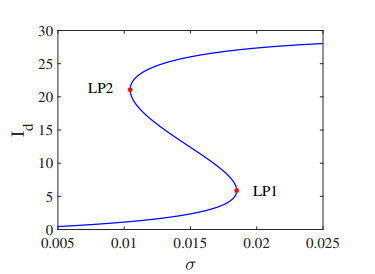
Testing-culling is a very effective measure for the prevention and control of animal diseases. In this paper, based on sheep brucellosis control policies and animal testing characteristics and considering the limitation of culling resources, a dynamic model is established to study the impact of testing-culling measure. Theoretical analysis reveals that the model may have one or three positive equilibria. The equilibrium in the middle is always unstable, and the model shows saddle-node bifurcation, generalized Hopf bifurcation and Bogdanov-Takens bifurcation. Moreover, the theoretical results are verified via numerical analysis. These results reveal that testing and culling strategies can induce complex transmission dynamics that can help us develop appropriate prevention and control measures for animal brucellosis.
Citation: Yongbing Nie, Xiangdong Sun, Hongping Hu, Qiang Hou. Bifurcation analysis of a sheep brucellosis model with testing and saturated culling rate[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 1519-1537. doi: 10.3934/mbe.2023069
Testing-culling is a very effective measure for the prevention and control of animal diseases. In this paper, based on sheep brucellosis control policies and animal testing characteristics and considering the limitation of culling resources, a dynamic model is established to study the impact of testing-culling measure. Theoretical analysis reveals that the model may have one or three positive equilibria. The equilibrium in the middle is always unstable, and the model shows saddle-node bifurcation, generalized Hopf bifurcation and Bogdanov-Takens bifurcation. Moreover, the theoretical results are verified via numerical analysis. These results reveal that testing and culling strategies can induce complex transmission dynamics that can help us develop appropriate prevention and control measures for animal brucellosis.

| [1] |
T. Barbier, F. Collard, A. Zúñiga-Ripa, I. Moriyón, T. Godard, J. Becker, et al., Erythritol feeds the pentose phosphate pathway via three new isomerases leading to D-erythrose-4-phosphate in Brucella, Proc. Natl. Acad. Sci. U. S. A., 111 (2014), 17815–17820. https://doi.org/10.1073/pnas.1414622111 doi: 10.1073/pnas.1414622111

|
| [2] |
P. Andriopoulos, M. Tsironi, S. Deftereos, A. Aessopos, G. Assimakopoulos, Acute brucellosis: Presentation, diagnosis, and treatment of 144 cases, Int. J. Infect. Dis., 11 (2007), 52–57. https://doi.org/10.1016/j.ijid.2005.10.011 doi: 10.1016/j.ijid.2005.10.011
|
| [3] |
G. Pappas, P. Papadimitriou, N. Akritidis, L. Christou, E. V. Tsianos, The new global map of human brucellosis, Lancet Infect. Dis., 6 (2006), 91–99. https://doi.org/10.1016/S1473-3099(06)70382-6 doi: 10.1016/S1473-3099(06)70382-6
|
| [4] |
H. Zeng, Y. M. Wang, X. D. Sun, P. Liu, Q. G. Xu, D. Huang, et al., Status and influencing factors of farmers' private investment in the prevention and control of sheep brucellosis in China: A cross-sectional study, PLoS Neglected Trop. Dis., 13 (2019), e0007285. https://doi.org/10.1371/journal.pntd.0007285 doi: 10.1371/journal.pntd.0007285
|
| [5] |
A. J. S. Alves, F. Rocha, M. Amaku, F. Ferreira, E. O. Telles, J. H. H. Grisi Filho, et al., Economic analysis of vaccination to control bovine brucellosis in the States of Sao Paulo and Mato Grosso, Brazil, Prev. Vet. Med., 118 (2015), 351–358. https://doi.org/10.1016/j.prevetmed.2014.12.010 doi: 10.1016/j.prevetmed.2014.12.010
|
| [6] |
Q. Hou, X. D. Sun, J. Zhang, Y. J. Liu, Y. M. Wang, Z. Jin, Modeling the transmission dynamics of sheep brucellosis in Inner Mongolia Autonomous Region, China, Math. Biosci., 242 (2013), 51–58. https://doi.org/10.1016/j.mbs.2012.11.012 doi: 10.1016/j.mbs.2012.11.012
|
| [7] |
H. Yoon, O. K. Moon, S. H. Lee, W. C. Lee, M. Her, W. Jeong, et al., Epidemiology of brucellosis among cattle in Korea from 2001 to 2011, J. Vet. Sci., 15 (2014), 537–543. https://doi.org/10.4142/jvs.2014.15.4.537 doi: 10.4142/jvs.2014.15.4.537
|
| [8] |
R. M. Davidson, Control and eradication of animal diseases in New Zealand, N. Z. Vet. J., 50 (2002), 6–12. https://doi.org/10.1080/00480169.2002.36259 doi: 10.1080/00480169.2002.36259
|
| [9] |
M. Zamri-Saad, M. I. Kamarudin, Control of animal brucellosis: The Malaysian experience, Asian Pac. J. Trop. Med., 9 (2016), 1136–1140. https://doi.org/10.1016/j.apjtm.2016.11.007 doi: 10.1016/j.apjtm.2016.11.007
|
| [10] |
N. Zhang, D. S. Huang, W. Wu, J. Liu, F. Liang, B. S. Zhou, et al., Animal brucellosis control or eradication programs worldwide: A systematic review of experiences and lessons learned, Prev. Vet. Med., 160 (2018), 105–115. https://doi.org/10.1016/j.prevetmed.2018.10.002 doi: 10.1016/j.prevetmed.2018.10.002
|
| [11] |
P. O. Lolika, C. Modnak, S. Mushayabasa, On the dynamics of brucellosis infection in bison population with vertical transmission and culling, Math. Biosci., 305 (2018), 42–54. https://doi.org/10.1016/j.mbs.2018.08.009 doi: 10.1016/j.mbs.2018.08.009
|
| [12] |
L. Bolzoni, R. D. Marca, M. Groppi, A. Gragnani, Dynamics of a metapopulation epidemic model with localized culling, Discrete Cont. Dyn.-B, 25 (2020), 2307–2330. https://doi.org/10.3934/dcdsb.2020036 doi: 10.3934/dcdsb.2020036
|
| [13] |
J. Zinsstag, F. Roth, D. Orkhon, G. Chimed-Ochir, M. Nansalmaa, J. Kolar, et al., A model of animal-human brucellosis transmission in Mongolia, Prev. Vet. Med., 69 (2005), 77–95. https://doi.org/10.1016/j.prevetmed.2005.01.017 doi: 10.1016/j.prevetmed.2005.01.017
|
| [14] |
M. F. Abakar, H. Yahyaoui Azami, P. Justus Bless, L. Crump, P. Lohmann, M. Laager, et al., Transmission dynamics and elimination potential of zoonotic tuberculosis in morocco, PLoS Neglected Trop. Dis., 11 (2017), e0005214. https://doi.org/10.1371/journal.pntd.0005214 doi: 10.1371/journal.pntd.0005214
|
| [15] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [16] |
G. H. Li, Y. X. Zhang, Dynamic behaviors of a modified SIR model in epidemic diseases using nonlinear incidence and recovery rates, PLoS One, 12 (2017), e0175789. https://doi.org/10.1371/journal.pone.0175789 doi: 10.1371/journal.pone.0175789
|
| [17] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, 3rd edition, Springer-Verlag, New York, 2004. https://doi.org/10.1007/978-1-4757-3978-7 |
| [18] |
D. A. Abernethy, J. Moscard-Costello, E. Dickson, R. Harwood, K. Burns, E. McKillop, et al., Epidemiology and management of a bovine brucellosis cluster in Northern Ireland, Prev. Vet. Med., 98 (2011), 223–229. https://doi.org/10.1016/j.prevetmed.2010.11.002 doi: 10.1016/j.prevetmed.2010.11.002
|



Figures(4)

Yongbing Nie, Xiangdong Sun, Hongping Hu, Qiang Hou. Bifurcation analysis of a sheep brucellosis model with testing and saturated culling rate[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 1519-1537. doi: 10.3934/mbe.2023069







 DownLoad:
DownLoad: