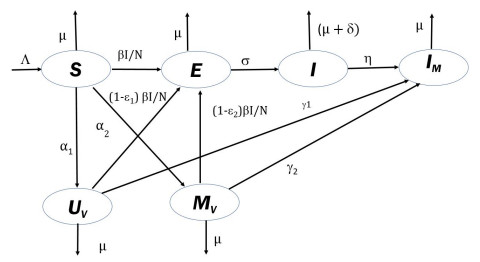
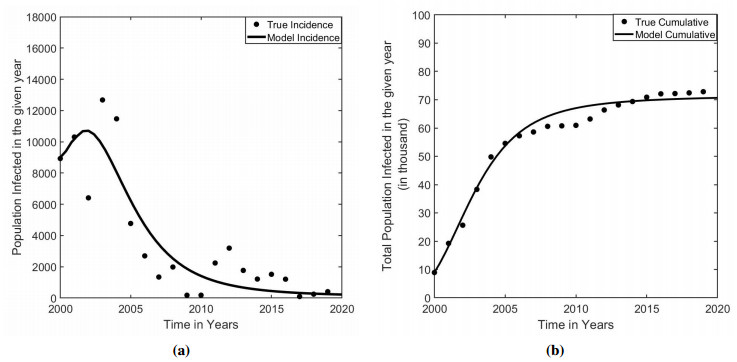
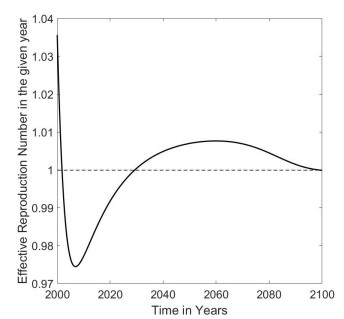
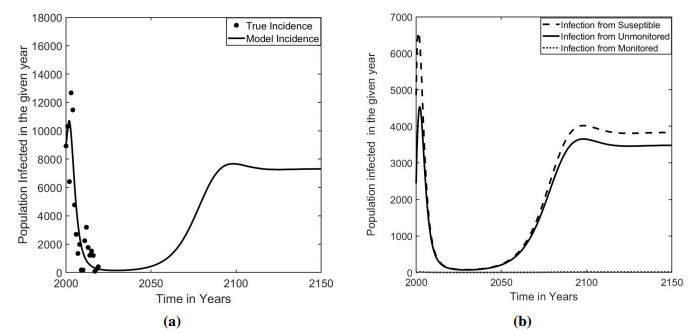
Measles is one of the highly contagious human viral diseases. Despite the availability of vaccines, measles outbreak frequently occurs in many places, including Nepal, partly due to the lack of compliance with vaccination. In this study, we develop a novel transmission dynamics model to evaluate the effects of monitored vaccination programs to control and eliminate measles. We use our model, parameterized with the data from the measles outbreak in Nepal, to calculate the vaccinated reproduction number, $ R_v $, of measles in Nepal. We perform model analyses to establish the global asymptotic stability of the disease-free equilibrium point for $ R_v < 1 $ and the uniform persistence of the disease for $ R_v > 1 $. Moreover, we perform model simulations to identify monitored vaccination strategies for the successful control of measles in Nepal. Our model predicts that the monitored vaccination programs can help control the potential resurgence of the disease.
Citation: Anjana Pokharel, Khagendra Adhikari, Ramesh Gautam, Kedar Nath Uprety, Naveen K. Vaidya. Modeling transmission dynamics of measles in Nepal and its control with monitored vaccination program[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8554-8579. doi: 10.3934/mbe.2022397
Measles is one of the highly contagious human viral diseases. Despite the availability of vaccines, measles outbreak frequently occurs in many places, including Nepal, partly due to the lack of compliance with vaccination. In this study, we develop a novel transmission dynamics model to evaluate the effects of monitored vaccination programs to control and eliminate measles. We use our model, parameterized with the data from the measles outbreak in Nepal, to calculate the vaccinated reproduction number, $ R_v $, of measles in Nepal. We perform model analyses to establish the global asymptotic stability of the disease-free equilibrium point for $ R_v < 1 $ and the uniform persistence of the disease for $ R_v > 1 $. Moreover, we perform model simulations to identify monitored vaccination strategies for the successful control of measles in Nepal. Our model predicts that the monitored vaccination programs can help control the potential resurgence of the disease.

| [1] | J. Hamborsky, A. Kroger, S. Wolfe, Epidemiology and prevention of vaccine-preventable diseases, 13th ed. Washington D.C. Public Health Foundation, 2015. |
| [2] |
E. Stephen, K. Raymond, K. Gabriel, F. Nestory, M. Godfrey, M. Arbogast, A mathematical model for control and elimination of the transmission dynamics of measles, Appl. Comput. Math., 4 (2015), 396–408. https://doi.org/10.11648/j.acm.20150406.12 doi: 10.11648/j.acm.20150406.12

|
| [3] | Center for Disease Control and Prevention (CDC), Measles epidemic: Overcoming vaccine prejudice (History of measles outbreaks), A Train Education, available from: https://www.atrainceu.com/content/2-history-and-pathology-measles (accessed on 8 February 2021). |
| [4] | World Health Organization, Measles, available from: https://www.who.int/news-room/fact-sheets/detail/measles, (accessed on 10 March 2021). |
| [5] | UNICEF, UNICEF for every child, Over 20 million children worldwide missed out on measles vaccine annually in past 8 years, creating a pathway to current global outbreaks, available from: https://www.unicef.org/nepal/press-releases/over-20-million-children-worldwide-missed-out-measles-vaccine-annually-\past-8-years (accessed on 28 March 2020). |
| [6] | United Nations: UN News, Measles cases hit 23-year high last year, killing 200,000 as vaccination stalls, WHO says, available from: https://news.un.org/en/story/2020/11/1077482 (accessed on 10 March 2020). |
| [7] |
M. K. Patel, J. L. Goodson, J. P. Alexander Jr, K. Kretsinger, S. V. Sodha, C. Steulet, et al., Progress toward regional measles elimination– worldwide, 2000–2019, Morb. Mortal. Wkly. Rep., 69 (2020), 1700. https://doi.org/10.15585/mmwr.mm6848a1 doi: 10.15585/mmwr.mm6848a1
|
| [8] | Government of Nepal, Ministry of Health and Population, National immunization programme, available from: https://www.mohp.gov.np/eng/program/child-health-services/nip (accessed on 28 March 2021). |
| [9] | L. H. Sun, B. Guarino, Anti-vaxxers target communities battling measles, available from: https://www.washingtonpost.com/national/health-science/antivaxxers-go-viral-in-communities-battling-measles/2019/05/20/a476417c-78d7-11e9-bd25-c989555e7766_story.html(accessed: 20 April 2020). |
| [10] | B. Zadrozny, E. Edwards, Anti-vaccine groups take dangerous online harassment into the real world, available from: https://www.nbcnews.com/health/kids-health/anti-vaccine-groups-take-dangerous-harassment-offline-real-world-n1096461 (accessed on 20 April 2020). |
| [11] | New York City (NYC) Health, Measles, available from: https://www1.nyc.gov/site/doh/health/health-topics/measles.page (accessed on 10 August 2020). |
| [12] | B. Guarino, L. H. Sun, New York anti-vaccine event attracts pro-vaccine protests amid measles outbreak, available from: https://www.washingtonpost.com/health/2019/06/05/brooklyn-anti-vaccine-event-attracts-pro-vaccine-protests-amid-measles\-outbreak/, (accessed on 20 April 2020). |
| [13] | Public Health Update, National immunization schedule, Nepal (Updated), available from: https://publichealthupdate.com/national-immunization-schedule-nepal/ (accessed on 10 March 2020). |
| [14] |
S. Khanal, T. R. Sedai, G. R. Choudary, J. N. Giri, R. Bohara, R. Pant, et al., Progress toward measles elimination — Nepal, 2007–2014, Morb. Mortal. Wkly. Rep., 65 (2016), 206–210. https://doi.org/10.15585/mmwr.mm6508a3 doi: 10.15585/mmwr.mm6508a3
|
| [15] | B. K. Suvedi, Twenty-five years of immunization program in Nepal, Kathmandu Univ. Med. J., 3 (2005), 4. |
| [16] | Government of Nepal, Ministry of Health and Population Child Health Division, Department of Health Services (DoHS), National immunization programme, available from: http://www.chd.gov.np (accessed on 10 March 2020). |
| [17] | Child Health Division, DoHS, MoHP, National immunization program, reaching every child, comprehensive multi-year plan 2068–2072 (2011–2016), MoHP, Nepal (2011). |
| [18] | A. Poudel, Routin immunization: How to Measles Transmission, (2014), available from: http://arjunmaske.blogspot.com/2014/09/how-to-measlestransmission.html (accessed on 12 August 2020). |
| [19] | A. Poudel, Measles outbreak in Morang, Dang and Kapilvastu raises concern, available from: https://kathmandupost.com/national/2019/04/30/measles-outbreak-in-morang-dang-and-kapilvastu-raises-concern (accessed on 12 September 2020). |
| [20] | A. Poudel, Low vaccine coverage rate, floating population leading to repeat measles outbreaks, available from: https://reliefweb.int/report/nepal/low-vaccine-coverage-rate-floating-population-leading-repeat-measles-outbreaks (accessed on 12 September 2020). |
| [21] | New Spotlight Online, Number Of Measles Cases In Nepal Increased by more than two times in 2018, available from: https://www.spotlightnepal.com/2019/04/25/number-measles-cases-nepal-increased/ (accessed on 25 March 2020). |
| [22] | A. Poudel, Measles outbreaks reported in five districts including in Kathmandu and Lalitpur in last one month, available from: https://kathmandupost.com/national/2020/04/29/measles-outbreaks-reported-in-five-districts-including-in-kathmandu-and-lalitpur\-in-last-one-month, (accessed on 5 March 2021). |
| [23] | S. Uprety, Meanwhile, a measles outbreak in Nepal, available from: https://www.nepalitimes.com/here-now/meanwhile-a-measles-outbreak-in-nepal/ (accessed on 28 March 2021). |
| [24] | K. B. Karki, M. Dhimal, A. R. Pandey, B. Bista, A. Pandey, B. R. Giri, et al., Measles outbreak in Kapilvastu, Nepal: an outbreak investigation, Nepal Health Research Council, Ramshah Path, Kathmandu, Nepal, 2016. |
| [25] | M. Poudel, Unidentified disease claims lives of six kids, available from: https://kathmandupost.com/national/2016/01/20/unidentified-disease-claims-lives-of-six-kids (accessed on 10 September 2020). |
| [26] |
S. Sitaula, G. R. Awasthi, J. B. Thapa, K. P Joshi, A. Ramaiya, Measles outbreak among unvaccinated children in Bajura, JNMA J. Nepal Med. Assoc., 50 (2010), 273–276. https://doi.org/10.31729/jnma.48 doi: 10.31729/jnma.48
|
| [27] | H. Trottier, P. Philippe, Deterministic modeling of infectious diseases: measles cycles and the role of births and vaccination, Internet J. Infect. Dis., 2 (2002), 8 page. |
| [28] | H. Trottier, P. Philippe, Deterministic modeling of infectious diseases: theory and methods, Internet J. Infect. Dis., 1 (2000), 3. |
| [29] |
S. N. Mitku, Mathematical modeling and simulation study for the control and transmission dynamics of measles, Am. J. Appl. Math., 5 (2017), 99–107. https://doi.org/10.11648/j.ajam.20170504.11 doi: 10.11648/j.ajam.20170504.11
|
| [30] |
M. G. Robert, M. I. Tobias, Predicting and preventing measles epidemics in New Zealand, Epidemiol. Infect., 124 (2000), 279–287. https://doi.org/10.11648/j.ajam.20170504.11 doi: 10.11648/j.ajam.20170504.11
|
| [31] |
A. Franceschetti, A. Pugliese, Threshold behaviour of a SIR epidemic model with age structure and immigration, J. Math. Biol., 57 (2008), 1–27. https://doi.org/10.1007/s00285-007-0143-1 doi: 10.1007/s00285-007-0143-1
|
| [32] | J. F. Christopher, S. Katriona, C. Spencer, C. M. Jose, G. Christopher, G. James, et al., Measles outbreak response decision-making under uncertainty: a retrospective analysis, J. R. Soc. Interface, 15 (2017), 20170575. http://doi.org/10.1098/rsif.2017.0575R |
| [33] |
A. A. Momoh, M. O. Ibrahim, I. J. Uwanta, S. B. Manga, Mathematical model for control of measles epidemiology, Inter. J. Pure Appl. Math., 67 (2013), 707–718. https://doi.org/10.12732/ijpam.v87i5.4 doi: 10.12732/ijpam.v87i5.4
|
| [34] | M. D. la Sens, A. Quesadas, A. Ibeas, R. Nistal, An observed-based vaccination law for a SEIR epidemic model, Int. J. Comput. Theory Eng., 4 (2012), 1–32. http://doi.org/10.7763/IJCTE.2012.V4.488 |
| [35] |
E. F. Doungmo Goufo, S. C. Oukouomi Noutchie, S. Mugisha, A fractional SEIR epidemic model for spatial and temporal spread of measles in metapopulations, Abstr. Appl. Anal., 2014 (2014), 6. https://doi.org/10.1155/2014/781028 doi: 10.1155/2014/781028
|
| [36] |
J. Ochoche, R. Gweryina, A mathematical model of measles with vaccination and two phases of infectiousness, J. Math., 10 (2019), 95–105. https://doi.org/10.9790/5728-101495105 doi: 10.9790/5728-101495105
|
| [37] |
E. M, Musyoki, R. Ndung'u, S. Osman, A mathematical model for the transmission of measles with passive immunity, Int. J. Res. Math. Stat. Sci., 6 (2019), 1–8. https://doi.org/10.26438/ijsrmss/v6i2.18 doi: 10.26438/ijsrmss/v6i2.18
|
| [38] |
M. Fred, J. Sigey, J. A. Okello, J. Okwoyo, G. J. Kang'ethe, Mathematical modeling on the control of measles by vaccination: case study of KISⅡ County Kenya, SIJ Trans. Comput. Sci. Eng. Appl. (CSEA), 2 (2015), 38–46. https://doi.org/10.9756/SIJCSEA/V2I4/0203150101 doi: 10.9756/SIJCSEA/V2I4/0203150101
|
| [39] | O. Christopher, A. I. Ibrahim, A. S. Timothy, Mathematical model of the dynamics of measles under the combined effect of vaccination and measles therapy, Int. J. Sci. Technol., 6 (2017), 118–173. |
| [40] | M. J. Wanjau, R. Titus, C. Isaac, Mathematical modeling of the transmission dynamics of measles under the effect of vaccination, IOSR J. Math., 15 (2019), 10–19. |
| [41] | R. Y. M'pika Massoukou, S. C. Oukouomi Noutchie, R. Guiem, Global dynamics of an SVEIR model with age-dependent vaccination, infection, and latency, Abstr. Appl. Anal., 2018 (2018), 1–21. https://doi.org/10.1155/2018/8479638 |
| [42] | T. Bakhtiar, Control policy mix in measles transmission dynamics using vaccination, therapy, and treatment, Int. J. Math. Math. Sci., 2020 (2020), 1–20. |
| [43] |
Z. Memon, S. Qureshi, B. R. Memon, Mathematical analysis for a new nonlinear measles epidemiological system using real incidence data from Pakistan, Eur. Phys. J. Plus, 135 (2020), 1–21. https://doi.org/10.1140/epjp/s13360-020-00392-x doi: 10.1140/epjp/s13360-020-00392-x
|
| [44] |
D. Aldila, D. Asrianti, A deterministic model of measles with imperfect vaccination and quarantine intervention, Int. Conf. Mathematics: Pure, Appl. Computat., 1218 (2019), 012044–012054. https://doi.org/10.1088/1742-6596/1218/1/012044 doi: 10.1088/1742-6596/1218/1/012044
|
| [45] | Centers for Disease Control and Prevention (CDC), Progress in measles control–Nepal, 2000-2006, MMWR Morb. Mortal Wkly Rep., 56 (2007), 1028–1031. |
| [46] |
A. T. Truong, M. N. Mulders, D. C. Gautam, W. Ammerlaan, R. L. de Swart, C. C. King, et al., Genetic analysis of Asian measles virus strains–new endemic genotype in Nepal, Virus Res., 76 (2001), 71–78. https://doi.org/10.1016/S0168-1702(01)00255-6 doi: 10.1016/S0168-1702(01)00255-6
|
| [47] |
A. B. Joshi, Measles deaths in Nepal: estimating the national cases in fatality ratio, Bull World Health Organ, 87 (2009), 405–484. https://doi.org/10.2471/BLT.07.050427 doi: 10.2471/BLT.07.050427
|
| [48] | World Health Organization, WHO vaccine-preventable diseases: monitoring system. 2020 global summary, 2019, available from: https://apps.who.int/immunization_monitoring/globalsummary (accessed on 28 march 2020). |
| [49] |
B. Pantha, S. Giri, H. R. Joshi, N. K.Vaidya, Modeling transmission dynamics of rabies in Nepal, Infect. Dis. Model., 6 (2021), 284–301. https://doi.org/10.1016/j.idm.2020.12.009 doi: 10.1016/j.idm.2020.12.009
|
| [50] | Macrotrends, Nepal Population 1950–2020, available from: https://www.macrotrends.net/countries/NPL/nepal/population (accessed on 2 October 2020). |
| [51] | World Health Organization, Nepal - Deaths due to measles among children aged < 5 years, available from: https://www.who.int/immunization/monitoring_surveillance/burden/vpd/WHO_SurveillanceVaccinePreventable_11_Measles_R1.pdf?ua=1 (accessed on 2 October 2020). |
| [52] | World Data Atlas, Nepal-Deaths due to measles among children aged < 5 years, available from: https://knoema.com/atlas/Nepal/topics/Health/Deaths-among-children-under-5-by-cause/Deaths-due-to-measles (accessed on 3 October 2020). |
| [53] | H. S. Rathore, Measles outbreak kills two children and infects over 150 in Chepang settlements of Dhading district, available from: https://kathmandupost.com/province-no-3/2020/04/06/measles-outbreak-kills-two-children-and-infects-over-150-in-chepang-\settlements-of-dhading-district (accessed on 3 October 2020). |
| [54] | H. Motulasky, A. Christopoulos, Fitting Model to Biological Data Using Linear and Non Linear Regression. A Practical Guide to Curve Fitting, Graph Pad Software Inc.San Diego, CA, (2003), www.graphpad.com https://doi.org/10.5206/mase/10847. |
| [55] | M. Rahman, K. Bekele-Maxwell, L. L. Cates, H. T. Banks, N. K. Vaidya, Modeling zika virus transmission dynamics: parameter estimates, disease characteristics, and prevention, Sci. Rep., 9 (2019), 10575. https://doi.org/10.1038/s41598-019-46218-4 |
| [56] | H. T. Banks, S. Hu, W. C. Tompson, Modeling and inverse problems in the presence of uncertainty, CRC Press, Taylor and Francis Group, A Chapman and Hall Book, 2014. |
| [57] |
H. T. Banks, M. L. Joyner, Information content in data sets: a review of methods for interrogation and model comparison, J. Inverse Ill-Posed Probl., 26 (2018), 423–452. https://doi.org/10.1515/jiip-2017-0096 doi: 10.1515/jiip-2017-0096
|
| [58] |
K. Adhikari, R. Gautam, A. Pokharel, K. N. Uprety, N. K. Vaidya, Transmission dynamics of COVID-19 in Nepal: Mathematical model uncovering effective control, J. Theor. Biol., 521 (2021), 110680. https://doi.org/10.1016/j.jtbi.2021.110680 doi: 10.1016/j.jtbi.2021.110680
|
| [59] | C. D. Schunn, D. P. Wallach, Evaluating goodness-of-fit in comparison of models to data, Psychologie der Kognition: Reden and Vorträge anlässlich der Emeritierung von Werner Tack, (2005), 115–135. |
| [60] |
H. Miao, X. Xia, A. S. Perelson, H. Wu, On identifiability of nonlinear ODE models and applications in viral dynamics, SIAM Rev., 53 (2011), 3–39. https://doi.org/10.1137/090757009 doi: 10.1137/090757009
|
| [61] | O. Diekmann, J. A. P. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, Wiley, New York, 1999. |
| [62] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [63] | G. Chowell, J. M. Hyman, Mathematical and statistical modeling for emerging and re-emerging infectious diseases, Springer International Publishing Switzerland, 2016. |
| [64] |
O. Diekmann, J. A. P. Heesterbeek, M. G. Roberts, The construction of next-generation matrices for compartmental epidemic models, J. R. Soc. Interface, 7 (2010), 873–885. https://doi.org/10.1098/rsif.2009.0386 doi: 10.1098/rsif.2009.0386
|
| [65] |
M. A. Kuddus, M. Mohiuddin, A. Rahman, Mathematical analysis of a measles transmission dynamics model in Bangladesh with double dose vaccination, Sci. Rep., 11 (2021), 1–16. https://doi.org/10.1038/s41598-021-95913-8 doi: 10.1038/s41598-021-95913-8
|
| [66] | H. L. Smith, P. Waltman, The theory of the chemostat, Cambridge University Press, 1995. |
| [67] |
J. M. Mutua, F. Bin Wang, N. K. Vaidya, Modeling malaria and typhoid fever co-infection dynamics, Math. Biosci., 264 (2015), 128–144. https://doi.org/10.1016/j.mbs.2015.03.014 doi: 10.1016/j.mbs.2015.03.014
|
| [68] | T. Kato, Perturbation theory for linear operators, Springer-Verlag, Berlin Heidelberg, 1976. |
| [69] | X. Q. Zhao, Dynamical systems in population biology, Springer International Publishing, 2017. |
| [70] | Centers for Disease Control and Prevention, Measles symptoms and complications, available from: https://www.cdc.gov/measles/symptoms/index.html (accessed on 10 September 2020). |
| [71] |
S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178–196. https://doi.org/10.1016/j.jtbi.2008.04.011 doi: 10.1016/j.jtbi.2008.04.011
|
| [72] |
J. Horrocks, C. T. Bauc, Algorithmic discovery of dynamic models from infectious disease data, Sci. Rep., 10 (2020), 7061. https://doi.org/10.1038/s41598-020-63877-w doi: 10.1038/s41598-020-63877-w
|
Figures(9) / Tables(1)

Anjana Pokharel, Khagendra Adhikari, Ramesh Gautam, Kedar Nath Uprety, Naveen K. Vaidya. Modeling transmission dynamics of measles in Nepal and its control with monitored vaccination program[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 8554-8579. doi: 10.3934/mbe.2022397







 DownLoad:
DownLoad: