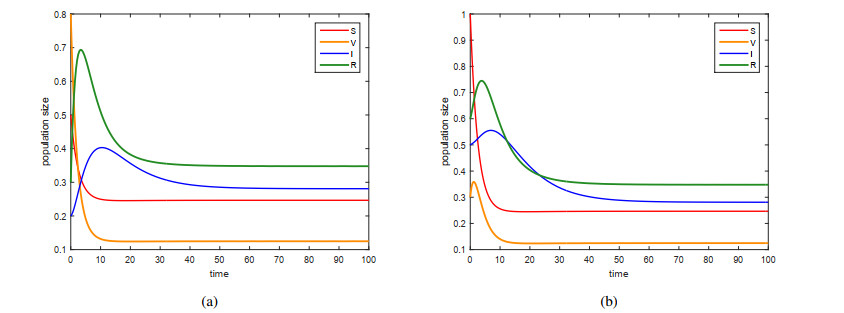
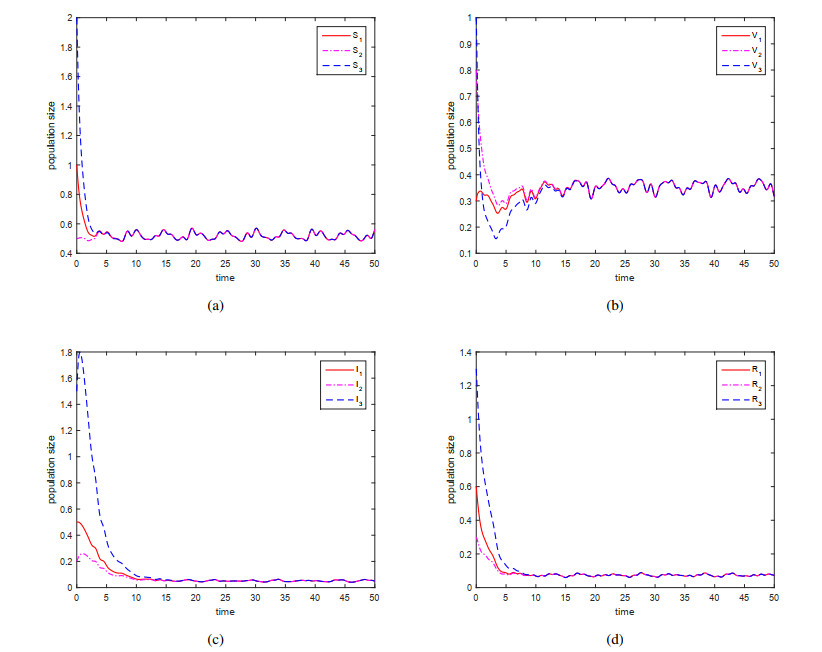
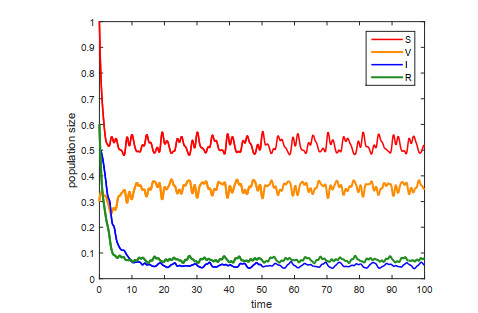
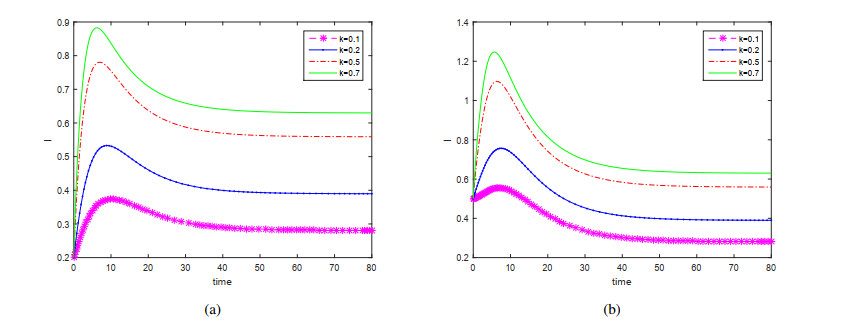
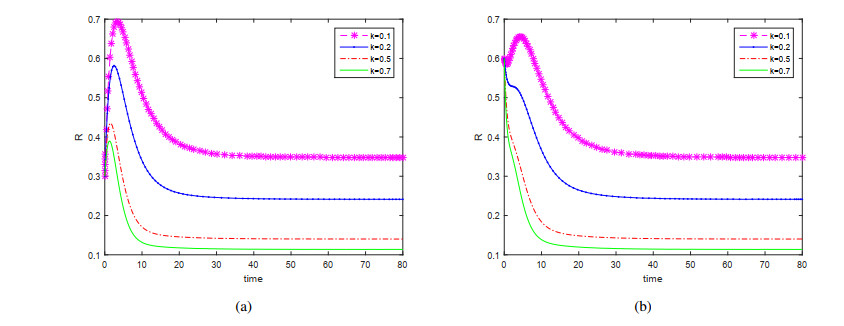
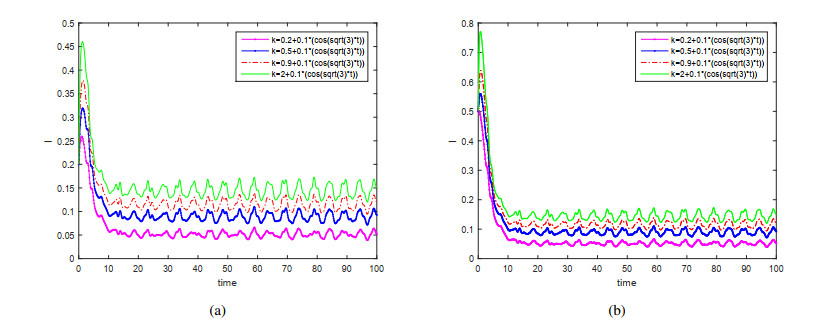
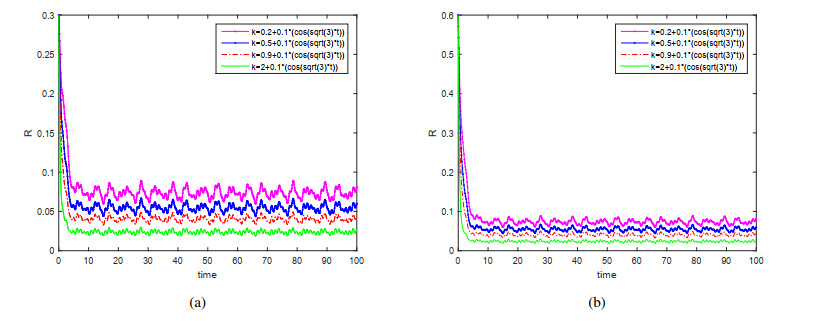
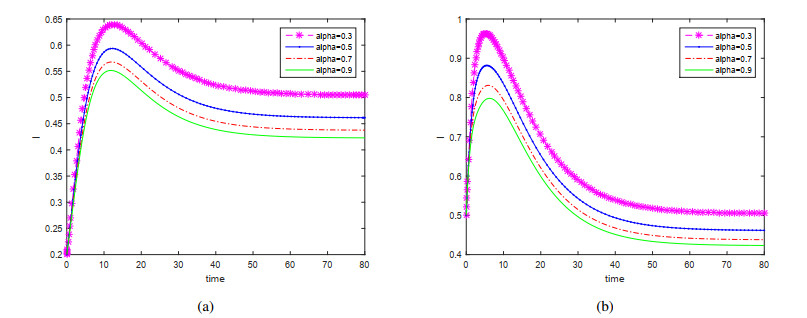
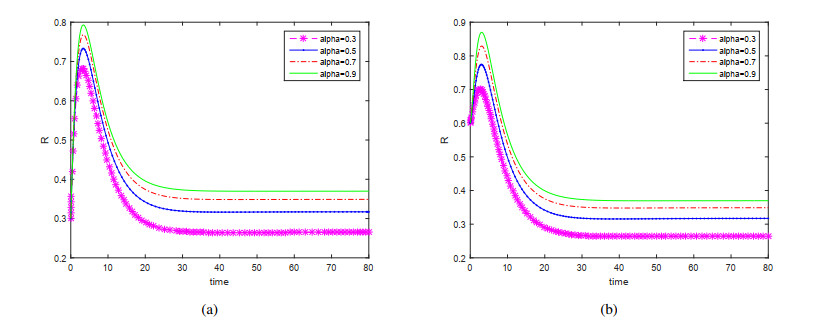
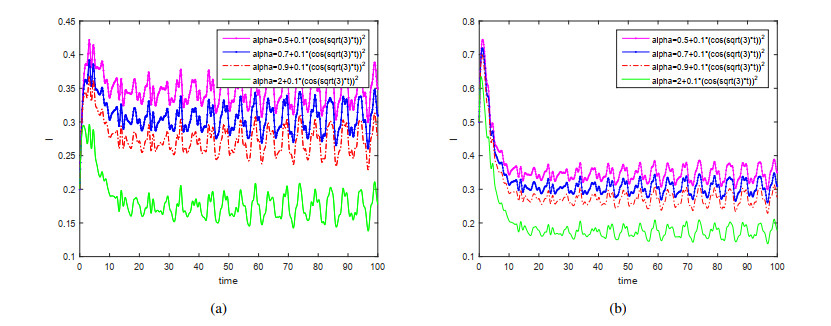
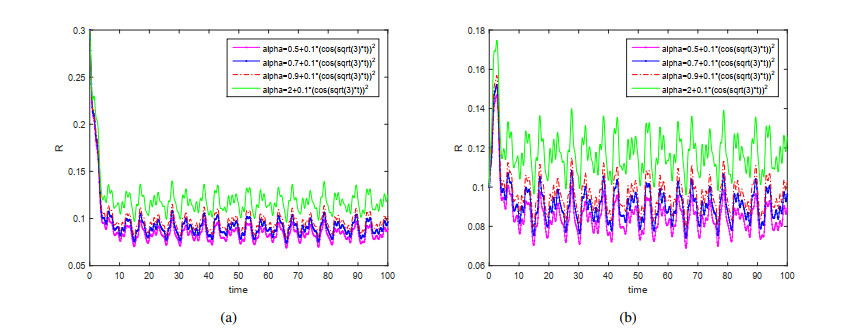
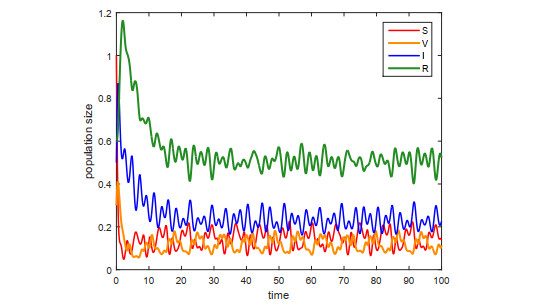
This paper is devoted to a nonautonomous SVIR epidemic model with relapse, that is, the recurrence rate is considered in the model. The permanent of the system is proved, and the result on the existence and uniqueness of globally attractive almost periodic solution of this system is obtained by constructing a suitable Lyapunov function. Some analysis for the necessity of considering the recurrence rate in the model is also presented. Moreover, some examples and numerical simulations are given to show the feasibility of our main results. Through numerical simulation, we have obtained the influence of vaccination rate and recurrence rate on the spread of the disease. The conclusion is that in order to control the epidemic of infectious diseases, we should increase the vaccination rate while reducing the recurrence rate of the disease.
Citation: Yifan Xing, Hong-Xu Li. Almost periodic solutions for a SVIR epidemic model with relapse[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 7191-7217. doi: 10.3934/mbe.2021356
This paper is devoted to a nonautonomous SVIR epidemic model with relapse, that is, the recurrence rate is considered in the model. The permanent of the system is proved, and the result on the existence and uniqueness of globally attractive almost periodic solution of this system is obtained by constructing a suitable Lyapunov function. Some analysis for the necessity of considering the recurrence rate in the model is also presented. Moreover, some examples and numerical simulations are given to show the feasibility of our main results. Through numerical simulation, we have obtained the influence of vaccination rate and recurrence rate on the spread of the disease. The conclusion is that in order to control the epidemic of infectious diseases, we should increase the vaccination rate while reducing the recurrence rate of the disease.

| [1] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. Lond. Ser. A., 115 (1927), 700-721. doi: 10.1098/rspa.1927.0118

|
| [2] | K. Fushimi, Y. Enatsu, E. Ishiwata, Global stability of an SIS epidemic model with delays, Math. Appl. Sci., 41 (2018), 5345-5354. |
| [3] |
H. A. A. El-Saka, The fractional-order SIS epidemic model with variable population size, Egyptian Math. Soc., 22 (2014), 50-54. doi: 10.1016/j.joems.2013.06.006
|
| [4] |
D. Bichara, Y. Kang, C. Castillo-Chavez, R. Horan, C. Perrings, SIS and SIR epidemic models under virtual dispersal, Bull. Math. Biol., 77 (2015), 2004-2034. doi: 10.1007/s11538-015-0113-5
|
| [5] |
T. Kuniya, J. Wang, Global dynamics of an SIR epidemic model with nonlocal diffusion, Nonlinear Anal. Real World Appl., 43 (2018), 262-282. doi: 10.1016/j.nonrwa.2018.03.001
|
| [6] | P. A. Naik, J. Zu, M. Ghoreishi, Stablity anaysis and approximate solution of sir epidemic model with crowley-martin type functional response and holling type-II treatment rate by using homotopy analysis method, J. Appl. Anal. Comput., 10 (2020), 1482-1515. |
| [7] | P. A. Naik, Global dynamics of a fractional-order SIR epidemic model with memory, Int. J. Biomath., 13 (2020), 1-23. |
| [8] |
C. N. Angstmann, B. I. Henry, A. V. Mcgann, A fractional order recovery SIR model from a stochastic process, Bull. Math. Biol., 78 (2016), 468-499. doi: 10.1007/s11538-016-0151-7
|
| [9] | P. A. Naik, J. Zu, K. M. Owolabi, Global dynamics of a fractional order model for the transmission of HIV epidemic with optimal control, Chaos Solitons Fractals, 138 (2020), 1-30. |
| [10] | P. A. Naik, K. M. Owolabi, M. Yavuz, J. Zu, Chaotic dynamics of a fractional order HIV-1 model involving aids-related cancer cells, Chaos Solitons Fractals, 140 (2020), 1-13. |
| [11] | P. A. Naik, J. Zu, K. M. Owolabi, Modeling the mechanics of viral kinetics under immune control during primary infection of HIV-1 with treatment in fractional order, Phys. A, 545 (2020), 1-19. |
| [12] |
P. A. Naik, M. Yavuz, S. Qureshi, J. Zu, S. Townley, Modeling and analysis of COVID-19 epidemics with treatment in fractional derivatives using real data from pakistan, Eur. Phys. J. Plus, 135 (2020), 795. doi: 10.1140/epjp/s13360-020-00819-5
|
| [13] | S. X. Zhang, H. Guo, Global analysis of age-structured multi-stage epidemic models for infectious diseases, Appl. Math. Comput., 337 (2018), 214-233. |
| [14] |
P. Liu, H. X. Li, Global behavior of a multi-group SEIR epidemic model with age structure and spatial diffusion, Math. Biosci. Eng., 17 (2020), 7248-7273. doi: 10.3934/mbe.2020372
|
| [15] | A. Ricardo, Analysis of a fractional SEIR model with treatment, Appl. Math. Lett., 84 (2016), 56-62. |
| [16] |
J. Li, Y. Yang, Y. Zhou, Global stability of an epidemic model with latent stage and vaccination, Nonlinear Anal. Real World Appl., 12 (2011), 2163-2173. doi: 10.1016/j.nonrwa.2010.12.030
|
| [17] | H. F Huo, L. X. Feng, Global stability of an epidemic model with incomplete treatment and vaccination, Discrete Dyn. Nat. Soc., 2012 (2012), 87-88. |
| [18] | M. Baniyaghoub, R. Gautam, Z. Shuai, P. van den Driessche, R. Ivanek, Reproduction numbers for infections with free-living pathogens growing in the environment, Discrete Dyn. Nat. Soc., 6 (2012), 923-940. |
| [19] |
T. Caraco, I. N. Wang, Free-living pathogens: life-history constraints and strain competition, J. Theoret. Biol., 250 (2008), 569-579. doi: 10.1016/j.jtbi.2007.10.029
|
| [20] |
S. A. Boone, C. P. Gerba, Significance of fomites in the spread of respiratory and enteric viral disease, Appl. Environ. Microb., 73 (2007), 1687-1696. doi: 10.1128/AEM.02051-06
|
| [21] |
A. Gabbuti, L. Roman$\grave{o}$, P. Blanc, F. Meacci, A. Amendola, A. Mele, et al., Long-term immunogenicity of hepatitis B vaccination in a cohort of italian healthy adolescents, Vaccine, 25 (2007), 3129-3132. doi: 10.1016/j.vaccine.2007.01.045
|
| [22] | F. Xu, R. Cressman, Disease control through voluntary vaccination decisions based on the smoothed best response, Comput. Math. Methods Med., 2014 (2014), 825734. |
| [23] |
F. Xu, R. Cressman, Voluntary vaccination strategy and the spread of sexually transmitted diseases, Math. Biosci., 274 (2016), 94-107. doi: 10.1016/j.mbs.2016.02.004
|
| [24] |
P. C. Jentsch, M. Anand, C. T. Bauch, Prioritising COVID-19 vaccination in changing social and epidemiological landscapes: a mathematical modelling study, Lancet Infect. Dis., 21 (2021), 1097-1106. doi: 10.1016/S1473-3099(21)00057-8
|
| [25] |
A. Ciccone, B. Zanotti, Covid-vaccination working group on cerebral venous thrombosis after, The importance of recognizing cerebral venous thrombosis following anti-COVID-19 vaccination, Eur. J. Intern. Med., 89 (2021), 115-117. doi: 10.1016/j.ejim.2021.05.006
|
| [26] |
J. Q. Li, Y. L. Yang, Y. C. Zhou, Global stability of an epidemic model with latent stage and vaccination, Nonlinear Anal. Real World Appl., 12 (2011), 2163-2173. doi: 10.1016/j.nonrwa.2010.12.030
|
| [27] | H. F. Huo, L. X. Feng, Global stability of an epidemic model with incomplete treatment and vaccination, Discrete Dyn. Nat. Soc., 2012 (2012), 87-88. |
| [28] |
X. N. Liu, Y. Takeuchi, S. Iwami, SVIR epidemic models with vaccination strategies, J. Theor. Biol., 253 (2008), 1-11. doi: 10.1016/j.jtbi.2007.10.014
|
| [29] | H. W. Hethcote, S. A. Levin, Periodicity in Epidemiological Models, Springer Berlin Heidelberg, Berlin, Heidelberg, 1989. |
| [30] |
J. P. Tripathi, S. Abbas, Global dynamics of autonomous and nonautonomous SI epidemic models with nonlinear incidence rate and feedback controls, Nonlinear Dyn., 86 (2016), 337-3351. doi: 10.1007/s11071-016-2892-0
|
| [31] |
D. Cheban, Z. X. Liu, Periodic, quasi-periodic, almost periodic, almost automorphic, birkhoff recurrent and poisson stable solutions for stochastic differential equations, J. Differ. Equ., 269 (2020), 3652-3685. doi: 10.1016/j.jde.2020.03.014
|
| [32] |
Z. Du, Y. Lv, Permanence and almost periodic solution of a lotka-volterra model with mutual interference and time delays, Appl. Math. Model., 37 (2013), 1054-1068. doi: 10.1016/j.apm.2012.03.022
|
| [33] | H. H. Wu, S. B. Yu, Permanence, extinction, and almost periodic solution of a nicholson's blowflies model with feedback control and time delay, Discrete Dyn. Nat. Soc., 2013 (2013), 1-9. |
| [34] |
N. H. Du, N. N. Nhu, Permanence and extinction for the stochastic SIR epidemic model, J. Differ. Equ., 269 (2020), 9619-9652. doi: 10.1016/j.jde.2020.06.049
|
| [35] |
F. Chen, On a nonlinear nonautonomous predator-prey model with diffusion and distributed delay, J. Comput. Appl. Math., 180 (2005), 33-49. doi: 10.1016/j.cam.2004.10.001
|
| [36] |
Y. Q. Li, T. W. Zhang, Existence and multiplicity of positive almost periodic solutions for a non-autonomous SIR epidemic model, Bull. Malays. Math. Sci. Soc., 39 (2016), 359-379. doi: 10.1007/s40840-015-0176-3
|
| [37] | P. Y. Liu, H. X. Li, Global stability of autonomous and nonautonomous hepatitis b virus models in patchy environment, J. Appl. Math. Comput., 10 (2020), 1771-1799. |
| [38] | A. M. Fink, Almost periodic differential equations, Springer-Verlag, New York, 1974. |
| [39] | C. Y. He, Almost Periodic Differential Equations, Higher Education Publishing House, Beijing, 1992. (in Chinese). |
| [40] |
F. Chen, Z. Li, Y. Huang, Note on the permanence of a competitive system with infinite delay and feedback controls, Nonlinear Anal. Real World Appl., 8 (2007), 680-687. doi: 10.1016/j.nonrwa.2006.02.006
|
| [41] | K. Gopalasamy, Stability and Oscillation in Delay Equation of Population Dynamics, Kluwer Academic Publishers, Dordrecht, 1992. |
| [42] | C. Zhang, N. J. Huang, D. O'Regan, Almost periodic solutions for a volterra model with mutual interference and holling type III functional response, Appl. Math. Comput., 225 (2013), 503-511. |
| [43] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [44] |
N. Chitnis, J. M. Hyman, J. M. Cushing, Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model, Bull. Math. Biol., 70 (2008), 1272-1296. doi: 10.1007/s11538-008-9299-0
|
Figures(12) / Tables(1)

Yifan Xing, Hong-Xu Li. Almost periodic solutions for a SVIR epidemic model with relapse[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 7191-7217. doi: 10.3934/mbe.2021356







 DownLoad:
DownLoad: