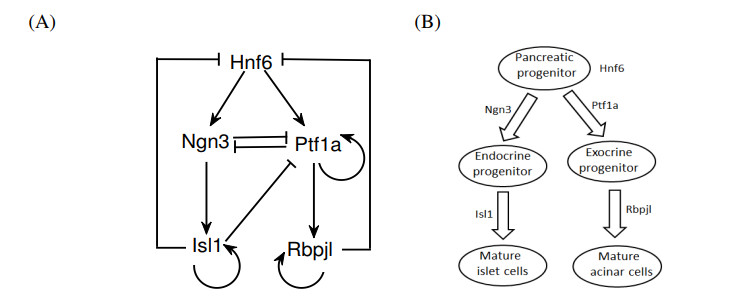
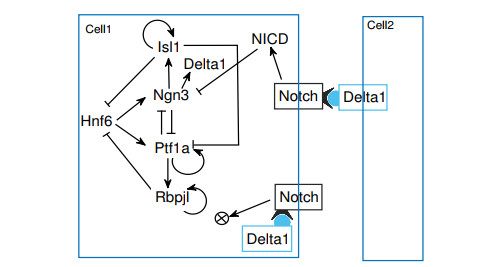
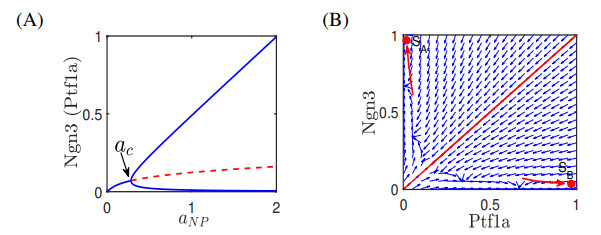
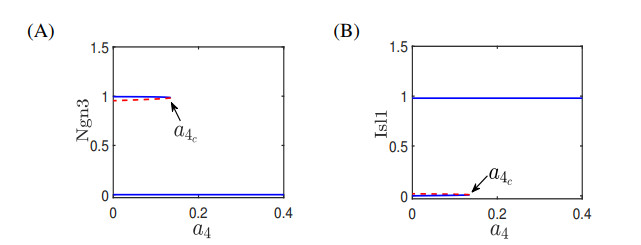
The endocrine and exocrine cells in pancreas originate initially from a group of apparently identical endoderm cells in the early gut. The endocrine and exocrine tissues are composed of islet/acinar and duct cells respectively. To explore the mechanism of pancreas cell fate decisions, we first construct a minimal mathematical model related to pancreatic regulations. The regulatory mechanism of acinar-to-islet cell conversion is revealed by bifurcation analysis of the model. In addition, Notch signaling is critical in determining the fate of endocrine and exocrine in the developing pancreas and it is a typical mediator of lateral inhibition which instructs adjacent cells to make different fate decisions. Next, we construct a multicellular model of cell-cell communication mediated by Notch signaling with trans-activation and cis-inhibition. The roles of Notch signaling in regulating fate decisions of endocrine and exocrine cells during the differentiation of pancreatic cells are explored. The results indicate that high (or low) level of Notch signaling drive cells to select the fate of exocrine (or endocrine) progenitor cells. The networks and the models presented here might be good candidates for providing qualitative mechanisms of pancreatic cell fate decisions. These results can also provide some insight on choosing perturbation strategies for further experimental analysis.
Citation: Dasong Huang, Ruiqi Wang. Exploring the mechanism of pancreatic cell fate decisions via cell-cell communication[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2401-2424. doi: 10.3934/mbe.2021122
The endocrine and exocrine cells in pancreas originate initially from a group of apparently identical endoderm cells in the early gut. The endocrine and exocrine tissues are composed of islet/acinar and duct cells respectively. To explore the mechanism of pancreas cell fate decisions, we first construct a minimal mathematical model related to pancreatic regulations. The regulatory mechanism of acinar-to-islet cell conversion is revealed by bifurcation analysis of the model. In addition, Notch signaling is critical in determining the fate of endocrine and exocrine in the developing pancreas and it is a typical mediator of lateral inhibition which instructs adjacent cells to make different fate decisions. Next, we construct a multicellular model of cell-cell communication mediated by Notch signaling with trans-activation and cis-inhibition. The roles of Notch signaling in regulating fate decisions of endocrine and exocrine cells during the differentiation of pancreatic cells are explored. The results indicate that high (or low) level of Notch signaling drive cells to select the fate of exocrine (or endocrine) progenitor cells. The networks and the models presented here might be good candidates for providing qualitative mechanisms of pancreatic cell fate decisions. These results can also provide some insight on choosing perturbation strategies for further experimental analysis.

| [1] | J. M. Slack, Developmental biology of the pancreas, Development, 121 (1995), 1569–1580. |
| [2] |
M. Sander, M. S. German, The beta cell transcription factors and development of the pancreas, J. Mol. Med., 75 (1997), 327–340. doi: 10.1007/s001090050118

|
| [3] |
H. Edlund, Developmental biology of the pancreas, Diabetes, 50 (2001), S5–S9. doi: 10.2337/diabetes.50.2007.S5
|
| [4] |
Z. Elisabetta, M. Anastasia, D. Nathalie, F. Alida, T. Natascia, M. Dirk, et al., Evolutionary conserved role of ptf1a in the specification of exocrine pancreatic fates, Dev. Biol., 268 (2004), 174–184. doi: 10.1016/j.ydbio.2003.12.016
|
| [5] |
E. Maria, D. S. Wilson, S. G. Michael, Gene expression cascades in pancreatic development, Mech. Develop., 120 (2003), 65–80. doi: 10.1016/S0925-4773(02)00333-7
|
| [6] |
S. K. Kim, R. J. MacDonald, Signaling and transcriptional control of pancreatic organogenesis, Curr. Opin. Genet. Dev., 12 (2002), 540–547. doi: 10.1016/S0959-437X(02)00338-6
|
| [7] |
H. Edlund, Organogenesis: pancreatic organogenesis developmental mechanisms and implications for therapy, Nat. Rev. Genet., 3 (2002), 524–532. doi: 10.1038/nrg841
|
| [8] | J. Patrick, M. D. Serge, J. Jan, G. Catherine, G. Gerard, G. François, et al., Transcription factor hepatocyte nuclear factor 6 regulates pancreatic endocrine cell differentiation and controls expression of the proendocrine gene $ngn3$, Mol. Cell. Biol., 20 (1997), 4445–4454. |
| [9] |
G. Gradwohl, A. Dierich, M. LeMeur, G. François, Neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas, PNAS, 97 (2000), 1607–1611. doi: 10.1073/pnas.97.4.1607
|
| [10] | V. M. Schwitzgebel, D. W. Scheel, J. R. Conners, J. Kalamaras, J. E. Lee, D. J. Anderson, et al., Expression of neurogenin3 reveals an islet cell precursor population in the pancreas, Development, 127 (2000), 3533–3542. |
| [11] | G. Gu, J. Dubauskaite, D. A. Melton, Direct evidence for the pancreatic lineage: $NGN3+$ cells are islet progenitors and are distinct from duct progenitors, Development, 129 (2002), 2447–2457. |
| [12] | A. Ulf, L. P. Samuel, M. J. Thomas, E. Thomas, E. Helena, Independent requirement for ISL1 in formation of pancreatic mesenchyme and islet cells, Nature, 385 (1997), 258–260. |
| [13] |
M. Cockell, B. J. Stevenson, M. Strubin, O. Hagenbüchle, P. K. Wellauer, Identification of a cell-specific DNA-binding activity that interacts with a transcriptional activator of genes expressed in the acinar pancreas, Mol. Cell. Biol., 9 (1989), 2464–2476. doi: 10.1128/MCB.9.6.2464
|
| [14] |
A. Krapp, M. Knöfler, S. Frutiger, G. J. Hughes, O. Hagenbuchle, P. K. Wellauer, The p48 DNA-binding subunit of transcription factor PTF1 is a new exocrine pancreas-specific basic helix-loop-helix protein, Embo. J., 15 (1996), 4317–4329. doi: 10.1002/j.1460-2075.1996.tb00806.x
|
| [15] |
J. X. Zhou, L. Brusch, S. Huang, Predicting pancreas cell fate decisions and reprogramming with a hierarchical multi-attractor model, PLoS ONE, 6 (2011), e14752. doi: 10.1371/journal.pone.0014752
|
| [16] |
H. Charlotte, D. Jonathan, S. Catherine, E. P. Christophe, Pancreatic acinar differentiation is guided by differential laminin deposition, Sci. Rep., 9 (2019), 2711. doi: 10.1038/s41598-019-39077-6
|
| [17] |
A. Smita, A. Colin, V. S. David, Computational models of the notch network elucidate mechanisms of context-dependent signaling, PLoS Comput. Biol., 5 (2009), e1000390. doi: 10.1371/journal.pcbi.1000390
|
| [18] |
A. Åsa, L. Hao, S. Lukas, B. Paul, J. A. David, H. Tasuku, et al., Notch signalling controls pancreatic cell differentiation, Nature, 400 (1999), 877–881. doi: 10.1038/23716
|
| [19] |
O. Junko, Y. Masato, M. Hideki, G. Tomokazu, N. Rika, M. Yuko, et al., p48 subunit of mouse PTF1 binds to RBP-Jk/CBF-1, the intracellular mediator of Notch signalling, and is expressed in the neural tube of early stage embryos, Genes Cells, 6 (2001), 345–360. doi: 10.1046/j.1365-2443.2001.00422.x
|
| [20] |
B. T. Hendrik, S. Elida, B. Johannes, K. H. P. Gerhard, H. A. Martin, Modeling coexistence of oscillation and Delta/Notch-mediated lateral inhibition in pancreas development and neurogenesis, J. Theor. Biol., 430 (2017), 32–44. doi: 10.1016/j.jtbi.2017.06.006
|
| [21] |
L. Baeyens, S. Bonné, T. Bos, I. Rooman, C. Peleman, T. Lahoutte, et al., Notch signaling as gatekeeper of rat acinar-to-beta-cell conversion in vitro, Gastroenterology, 136 (2009), 1750–1760. doi: 10.1053/j.gastro.2009.01.047
|
| [22] | B. Walter, Z. Roland, B. Lutz, Transdifferentiation of pancreatic cells by loss of contact-mediated signaling, BMC Syst. Bio., 7 (2013). |
| [23] |
S. Marika, R. A. Emma, Do as I say, Not(ch) as I do: Lateral control of cell fate, Dev. Biol., 447 (2019), 58–70. doi: 10.1016/j.ydbio.2017.09.032
|
| [24] |
R. Wang, K. Liu, L. Chen, K. Aihara, Neural fate decisions mediated by trans-activation and cis-inhibition in Notch signaling, Bioinformatics, 27 (2011), 3158–3165. doi: 10.1093/bioinformatics/btr551
|
| [25] |
A. C. Miller, E. L. Lyons, T. G. Herman, Cis-inhibition of Notch by endogenous delta biases the outcome of lateral inhibition, Curr. Biol., 19 (2009), 1378–1383. doi: 10.1016/j.cub.2009.06.042
|
| [26] |
R. Kageyama, T. Ohtsuka, H. Shimojo, I. Imayoshi, Dynamic Notch signaling in neural progenitor cells and a revised view of lateral inhibition, Nat. Neurosci., 11 (2008), 1247–1251. doi: 10.1038/nn.2208
|
| [27] |
K. Uriu, Y. Morishita, Y. Iwasa, Synchronized oscillation of the segmentation clock gene in vertebrate development, J. Math. Biol., 61 (2010), 207–229. doi: 10.1007/s00285-009-0296-1
|
| [28] |
S. Huang, Y. P. Guo, G. May, T. Enver, Bifurcation dynamics in lineage-commitment in bipotent progenitor cells, Dev. Biol., 305 (2007), 695–713. doi: 10.1016/j.ydbio.2007.02.036
|
| [29] |
B. D. Macarthur, A. Maayan, I. R. Lemischka, Systems biology of stem cell fate and cellular reprogramming, Nat. Rev. Mol. Cell Biol., 10 (2009), 672–681. doi: 10.1038/nrm2766
|
| [30] |
M. Toshihiko, H. S. Galvin, A. H. Michael, M. M. David, E. J. Jane, J. M. Raymond, Transcriptional autoregulation controls pancreatic Ptf1a expression during development and adulthood, Mol. Cell. Biol., 28 (2008), 5458–5468. doi: 10.1128/MCB.00549-08
|
| [31] |
M. Xavier, C. V. Eva, F. Marta, M. G. Ana, Á. O. Maria, C. U. Elena, et al., Gene expression dynamics after murine pancreatitis unveils novel roles for Hnf1$\alpha$ in acinar cell homeostasis, Gut, 61 (2012), 1187–1196. doi: 10.1136/gutjnl-2011-300360
|
| [32] |
A. H. Efsun, M. B. Cecil, K. K. Seung, Gene regulatory networks governing pancreas development, Dev. Cell, 25 (2013), 5–13. doi: 10.1016/j.devcel.2013.03.016
|
| [33] |
J. Jensen, Gene regulatory factors in pancreatic development, Dev. Dynam., 229 (2004), 176–200. doi: 10.1002/dvdy.10460
|
| [34] |
A. Stathopoulos, M. Levine, Genomic regulatory networks and animal development, Dev. Cell, 9 (2005), 449–462. doi: 10.1016/j.devcel.2005.09.005
|
| [35] |
S. J. Tapscott, The circuitry of a master switch: Myod and the regulation of skeletal muscle gene transcription, Development, 132 (2005), 2685–2695. doi: 10.1242/dev.01874
|
| [36] |
M. Toshihiko, L. Qiaoming, M. B. Thomas, A. M. Mark, J. M. Raymond, Early pancreatic development requires the vertebrate Suppressor of Hairless (RBPJ) in the PTF1 bHLH complex, Gene. Dev., 21 (2007), 2629–2643. doi: 10.1101/gad.1575207
|
| [37] |
J. Mei, A. P. Ana, G. D. Tye, Q. H. Chinh, D. Daniel, A. H. David, et al., MIST1 and PTF1 collaborate in feed-forward regulatory loops that maintain the pancreatic acinar phenotype in adult mice, Mol. Cell. Biol., 36 (2016), 2945–2955. doi: 10.1128/MCB.00370-16
|
| [38] |
M. Stadtfeld, K. Hochedlinger, Induced pluripotency: history, mechanisms, and applications, Genes Dev., 24 (2010), 2239–2263. doi: 10.1101/gad.1963910
|
| [39] |
K. Plath, W. E. Lowry, Progress in understanding reprogramming to the induced pluripotent state, Nat. Rev. Genet., 12 (2011), 253–265. doi: 10.1038/nrg2955
|
| [40] |
I. Sancho-Martinez, S. H. Baek, B. J. C. Izpisua, Lineage conversion methodologiesmeet the reprogramming toolbox, Nat. Cell Biol., 14 (2012), 892–899. doi: 10.1038/ncb2567
|
| [41] |
Q. Zhou, J. Brown, A. Kanarek, J. Rajagopal, D. A. Melton, In vivo reprogramming of adult pancreatic exocrine cells to beta-cells, Nature, 455 (2008), 627–632. doi: 10.1038/nature07314
|
| [42] |
K. H. Song, S. H. Ko, Y. B. Ahn, S. J. Yoo, H. Chin, H. Kaneto, et al., In vitro transdifferentiation of adult pancreatic acinar cells into insulin-expressing cells, Biochem. Biophys. Res. Commun., 316 (2004), 1094–1100. doi: 10.1016/j.bbrc.2004.02.153
|
| [43] |
L. Baeyens, S. D. Breuck, J. Lardon, J. K. Mfopou, I. Rooman, L. Bouwens, In vitro generation of insulin-producing beta cells from adult exocrine pancreatic cells, Diabetologia, 48 (2005), 49–57. doi: 10.1007/s00125-004-1606-1
|
| [44] |
B. Walter, X. Z. Joseph, B. Lutz, On the role of lateral stabilization during early patterning in the pancreas, J. R. Soc. Interface, 10 (2013), 20120766. doi: 10.1098/rsif.2012.0766
|
| [45] |
C. L. Pin, J. M. Rukstalis, C. Johnson, S. F. Konieczny, The bHLH transcription factor Mist1 is required to maintain exocrine pancreas cell organization and acinar cell identity, J. Cell Biol., 155 (2001), 519–530. doi: 10.1083/jcb.200105060
|
| [46] |
J. Jan, E. P. Erna, G. Philip, H. Jacob, R. S. Heller, I. Makoto, et al., Control of endodermal endocrine development by Hes-1, Nat. Genet., 24 (2000), 36–44. doi: 10.1038/71657
|
| [47] |
E. Farzad, G. Bidyut, V. B. Andrew, W. L. John, A. A. Megan, Y. Xiaobing, et al., Notch inhibits Ptf1 function and acinar cell differentiation in developing mouse and zebrafish pancreas, Development, 131 (2004), 4213–4224. doi: 10.1242/dev.01280
|
| [48] |
S. David, L. Amit, L. Lauren, A. S. Leah, E. F. Michelle, A. A. Graham, et al., Cis-interactions between Notch and Delta generate mutually exclusive signalling states, Nature, 465 (2010), 86–90. doi: 10.1038/nature08959
|
| [49] |
S. Afelik, J. Jensen, Notch signaling in the pancreas: patterning and cell fate specification, WIREs Dev. Biol., 2 (2013), 531–544. doi: 10.1002/wdev.99
|
| [50] |
S. S. Senthil, Y. G. Chun, S. Z. Peggy, Notch signaling is required for lateral induction of Jagged1 during FGF-induced lens fiber differentiation, Dev. Biol., 332 (2009), 166–176. doi: 10.1016/j.ydbio.2009.05.566
|
| [51] |
X. Li, H. Chen, Z. Liu, Z. Ye, S. Gou, C. Wang, Overexpression of MIST1 reverses the epithelial-mesenchymal transition and reduces the tumorigenicity of pancreatic cancer cells via the Snail/Ecadherin pathway, Cancer Lett., 431 (2018), 96–104. doi: 10.1016/j.canlet.2018.05.043
|
| [52] |
A. Jonas, C. J. Mette, K. Rasmus, N. J. Jan, F. E. Martin, D. Tye, et al., Ptf1a control of Dll1 reveals an alternative to the lateral inhibition mechanism, Development, 139 (2012), 4492. doi: 10.1242/dev.090597
|
| [53] |
R. Benedito, M. Hellstr$\ddot{o}$m, Notch as a hub for signaling in angiogenesis, Exp. Cell Res., 319 (2013), 1281–1288. doi: 10.1016/j.yexcr.2013.01.010
|
| [54] |
B. Appel, L. A. Givan, J. S. Eisen, Delta-Notch signaling and lateral inhibition in zebrafish spinal cord development, BMC Dev. Biol., 1 (2001), 13. doi: 10.1186/1471-213X-1-13
|
| [55] |
P. Huang, F. Xiong, S. G. Megason, A. F. Schier, Attenuation of Notch and hedgehog signaling is required for fate specification in the spinal cord, PLoS Genet., 8 (2012), e1002762. doi: 10.1371/journal.pgen.1002762
|
| [56] |
P. P. Pr$\acute{e}$vot, C. Augereau, A. Simion, G. V. Steen, N. Dauguet, F. P. Lemaigre, P. Jacquemin, Let-7b and miR-495 stimulate differentiation and prevent metaplasia of pancreatic acinar cells by repressing HNF6, Gastroenterology, 145 (2013), 668–678. doi: 10.1053/j.gastro.2013.05.016
|
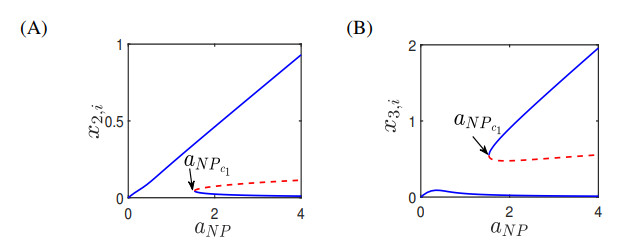
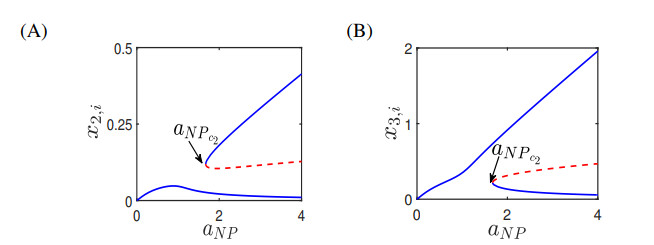
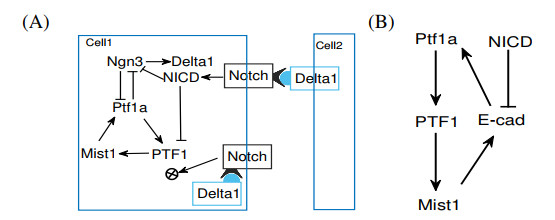
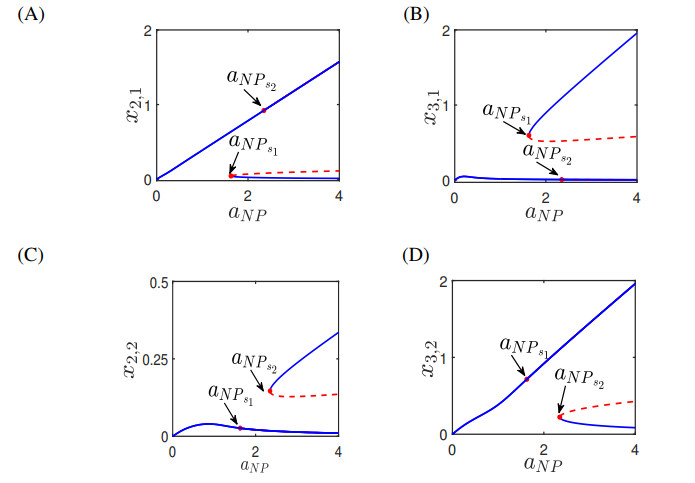
Figures(13) / Tables(4)

Dasong Huang, Ruiqi Wang. Exploring the mechanism of pancreatic cell fate decisions via cell-cell communication[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2401-2424. doi: 10.3934/mbe.2021122







 DownLoad:
DownLoad: