In order to treat the diseases caused by hepatitis C virus (HCV) more efficiently, the concentration of HCV in blood, cells, tissues and the body has attracted widespread attention from related scholars. This paper studies a dynamic dependent HCV model (more specifically, including age structure and treatment methods model) that concludes states of infection-free and infected equilibrium. Through eigenvalue analysis and Volterra integral formula, it proves that $ E_0 $ is globally asymptotically stable when $ \mathcal{R} < 1 $. After explaining the existence, uniqueness and positive properties of the solution of the system, we have proved the global asymptotic stability of $ E^* $ when $ \mathcal{R} > 1 $ by constructing a suitable Lyapunov function. Through the above proofs, it can be concluded that effective treatment measures can significantly reduce the number of HCVs, so many related researchers are aware of the importance of highly efficient nursing methods and are committed to applying relevant methods to practice.
Citation: Xiaowen Xiong, Yanqiu Li, Bingliang Li. Global stability of age-of-infection multiscale HCV model with therapy[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2182-2205. doi: 10.3934/mbe.2021110
In order to treat the diseases caused by hepatitis C virus (HCV) more efficiently, the concentration of HCV in blood, cells, tissues and the body has attracted widespread attention from related scholars. This paper studies a dynamic dependent HCV model (more specifically, including age structure and treatment methods model) that concludes states of infection-free and infected equilibrium. Through eigenvalue analysis and Volterra integral formula, it proves that $ E_0 $ is globally asymptotically stable when $ \mathcal{R} < 1 $. After explaining the existence, uniqueness and positive properties of the solution of the system, we have proved the global asymptotic stability of $ E^* $ when $ \mathcal{R} > 1 $ by constructing a suitable Lyapunov function. Through the above proofs, it can be concluded that effective treatment measures can significantly reduce the number of HCVs, so many related researchers are aware of the importance of highly efficient nursing methods and are committed to applying relevant methods to practice.

| [1] |
J. J. Feld, J. H. Hoofnagle, Mechanism of action of interferon and ribavirin in treatment of hepatitis C, Nature, 436 (2005), 967–972. doi: 10.1038/nature04082

|
| [2] |
B. Hajarizadeh, J. Grebely, H. McManus, C. Estes, H. Razavi, R. T. Gray, et al., Chronic hepatitis C burden and care cascade in australia in the era of interferon-based treatment, J. Gastroenterol. Hepatol., 32 (2017), 229–236. doi: 10.1111/jgh.13453
|
| [3] |
R. L. Morgan, B. Baack, B. D. Smith, A. Yartel, M. Pitasi, Y. Falck-Ytter, Eradication of hepatitis C virus infection and the development of hepatocellular carcinoma: a meta-analysis of observational studies, Ann. Intern. Med., 158 (2013), 329–337. doi: 10.7326/0003-4819-158-5-201303050-00005
|
| [4] |
P. Nahon, V. Bourcier, R. Layese, E. Audureau, C. Cagnot, P. Marcellin, et al., Eradication of hepatitis C virus infection in patients with cirrhosis reduces risk of liver and non-liver complications, Gastroenterology, 152 (2017), 142–156. doi: 10.1053/j.gastro.2016.09.009
|
| [5] |
T. Asselah, N. Boyer, D. Saadoun, M. Martinot-Peignoux, P. Marcellin, Direct-acting antivirals for the treatment of hepatitis C virus infection: optimizing current ifn-free treatment and future perspectives, Liver Int., 36 (2016), 47–57. doi: 10.1111/liv.13027
|
| [6] |
T. M. Nápoles, A. W. Batchelder, A. Lin, L. Moran, M. O. Johnson, M. Shumway, et al., Hcv treatment barriers among hiv/hcv co-infected patients in the us: a qualitative study to understand low uptake among marginalized populations in the daa era, J. Publ. Health, 41 (2019), e283–e289. doi: 10.1093/pubmed/fdz045
|
| [7] |
A. U. Neumann, N. P. Lam, H. Dahari, D. R. Gretch, T. E. Wiley, T. J. Layden, et al., Hepatitis c viral dynamics in vivo and the antiviral efficacy of interferon-$\alpha$ therapy, Science, 282 (1998), 103–107. doi: 10.1126/science.282.5386.103
|
| [8] |
J. Guedj, H. Dahari, L. Rong, N. D. Sansone, R. E. Nettles, S. J. Cotler, et al., Modeling shows that the ns5a inhibitor daclatasvir has two modes of action and yields a shorter estimate of the hepatitis C virus half-life, Proc. Natl. Acad. Sci., 110 (2013), 3991–3996. doi: 10.1073/pnas.1203110110
|
| [9] |
L. Rong, A. S. Perelson, Mathematical analysis of multiscale models for hepatitis c virus dynamics under therapy with direct-acting antiviral agents, Math. Biosci., 245 (2013), 22–30. doi: 10.1016/j.mbs.2013.04.012
|
| [10] |
L. Rong, J. Guedj, H. Dahari, D. J. Coffield Jr, M. Levi, P. Smith, et al., Analysis of hepatitis c virus decline during treatment with the protease inhibitor danoprevir using a multiscale model, PLoS Comput. Biol., 9 (2013), e1002959. doi: 10.1371/journal.pcbi.1002959
|
| [11] |
A. Goyal, Y. Lurie, E. G. Meissner, M. Major, N. Sansone, S. L. Uprichard, et al., Modeling hcv cure after an ultra-short duration of therapy with direct acting agents, Antiviral Res., 144 (2017), 281–285. doi: 10.1016/j.antiviral.2017.06.019
|
| [12] | B. M. Quintela, J. M. Conway, J. Hyman, R. Reis, R. dos Santos, M. Lobosco, et al., An age-based multiscale mathematical model of the hepatitis C virus life-cycle during infection and therapy: including translation and replication, Ⅶ Latin American Congress on Biomedical Engineering CLAIB 2016, 2016. |
| [13] | D. Echevarria, A. Gutfraind, B. Boodram, S. Cotler, M. Major, H. Dahari, Modeling vaccine and daa treatment scale up effect on hepatitis c prevalence among persons who inject drugs in metropolitan chicago, J. Hepatol., 66 (2017), S505–S506. |
| [14] | S. D. Lombardo, S. Lombardo, Global stability for a mathematical model of hepatitis c: Impact of a possible vaccination with daas therapy, AIP Conference Proceedings, 2019. |
| [15] |
K. Kitagawa, S. Nakaoka, Y. Asai, K. Watashi, S. Iwami, A pde multiscale model of hepatitis c virus infection can be transformed to a system of odes, J. Theor. Biol., 448 (2018), 80–85. doi: 10.1016/j.jtbi.2018.04.006
|
| [16] | H. L. Smith, H. R. Thieme, Dynamical systems and population persistence, American Mathematical Society, 2011. |
| [17] |
B. Soufiane, T. M. Touaoula, Global analysis of an infection age model with a class of nonlinear incidence rates, J. Math. Anal. Appl., 434 (2016), 1211–1239. doi: 10.1016/j.jmaa.2015.09.066
|
| [18] |
A. Chekroun, M. N. Frioui, T. Kuniya, T. M. Touaoula, Global stability of an age-structured epidemic model with general lyapunov functional, Math. Biosci. Eng., 16 (2019), 1525–1553. doi: 10.3934/mbe.2019073
|
| [19] |
M. N. Frioui, T. M. Touaoula, B. Ainseba, Global dynamics of an age-structured model with relapse, Discrete Contin. Dyna. Syst. B, 25 (2020), 2245. doi: 10.3934/dcdsb.2019226
|
| [20] | P. Magal, S. Ruan, Theory and applications of abstract semilinear Cauchy problems, Springer, 2018. |
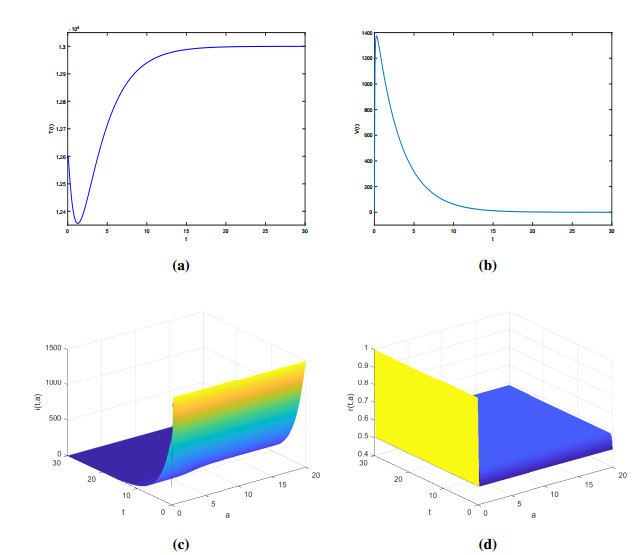
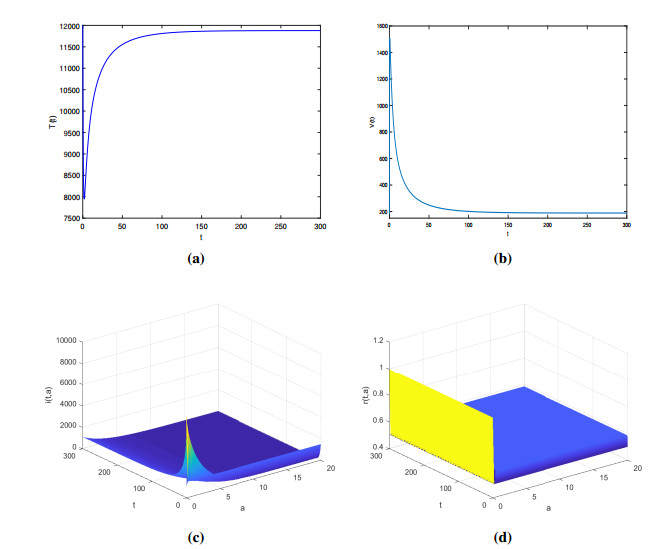
Figures(2)

Xiaowen Xiong, Yanqiu Li, Bingliang Li. Global stability of age-of-infection multiscale HCV model with therapy[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2182-2205. doi: 10.3934/mbe.2021110







 DownLoad:
DownLoad: